

A leptin-regulated circuit controls glucose mobilization during noxious stimuli
- PMID: 28714862
- PMCID: PMC5531403
- DOI: 10.1172/JCI90147
A leptin-regulated circuit controls glucose mobilization during noxious stimuli
Abstract
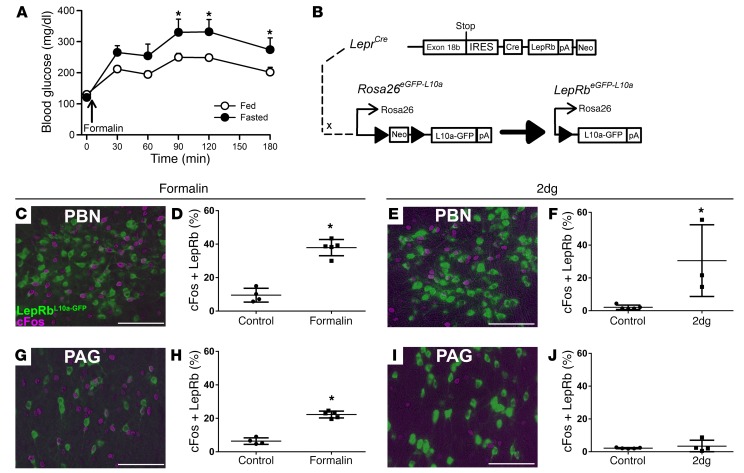
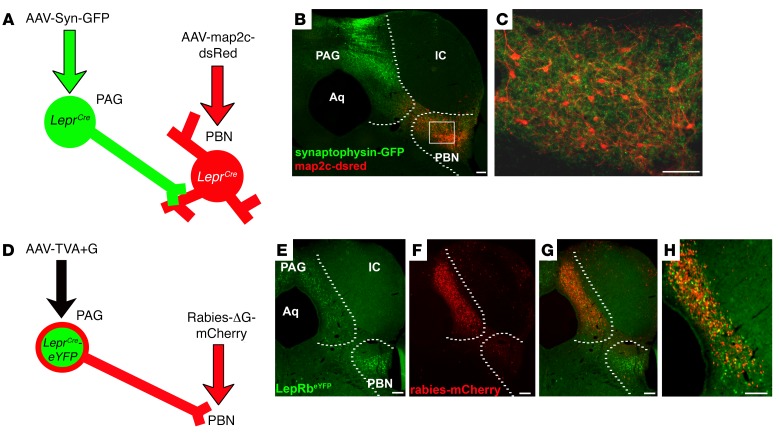
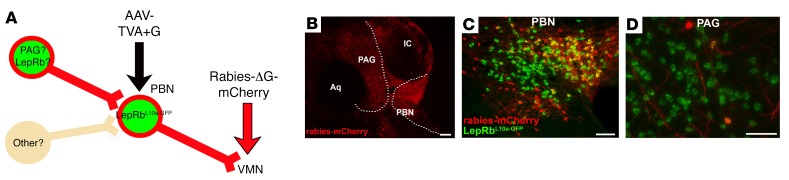
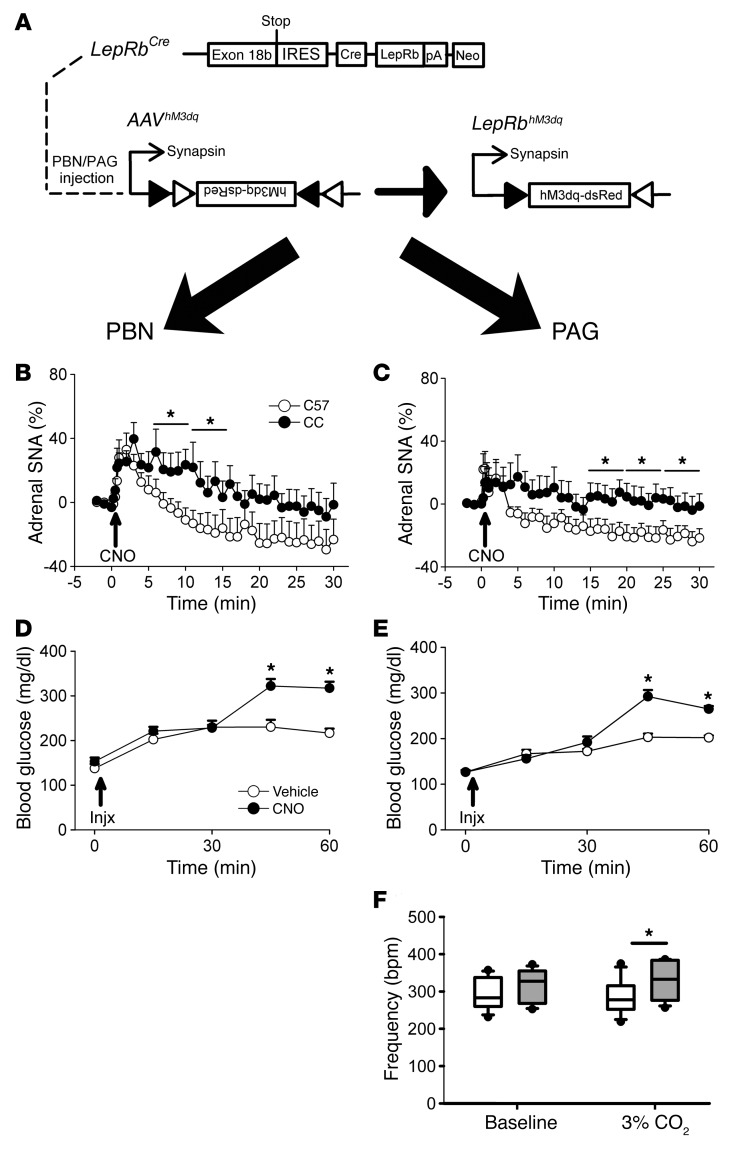
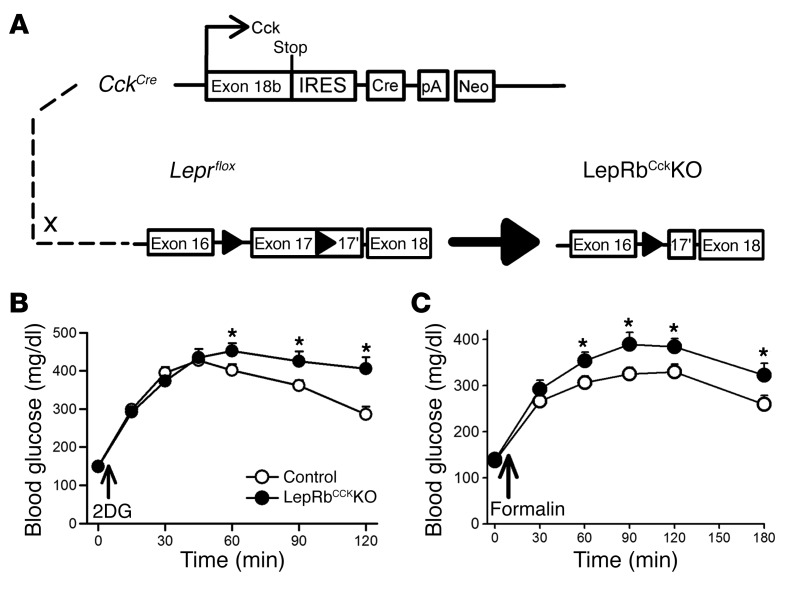
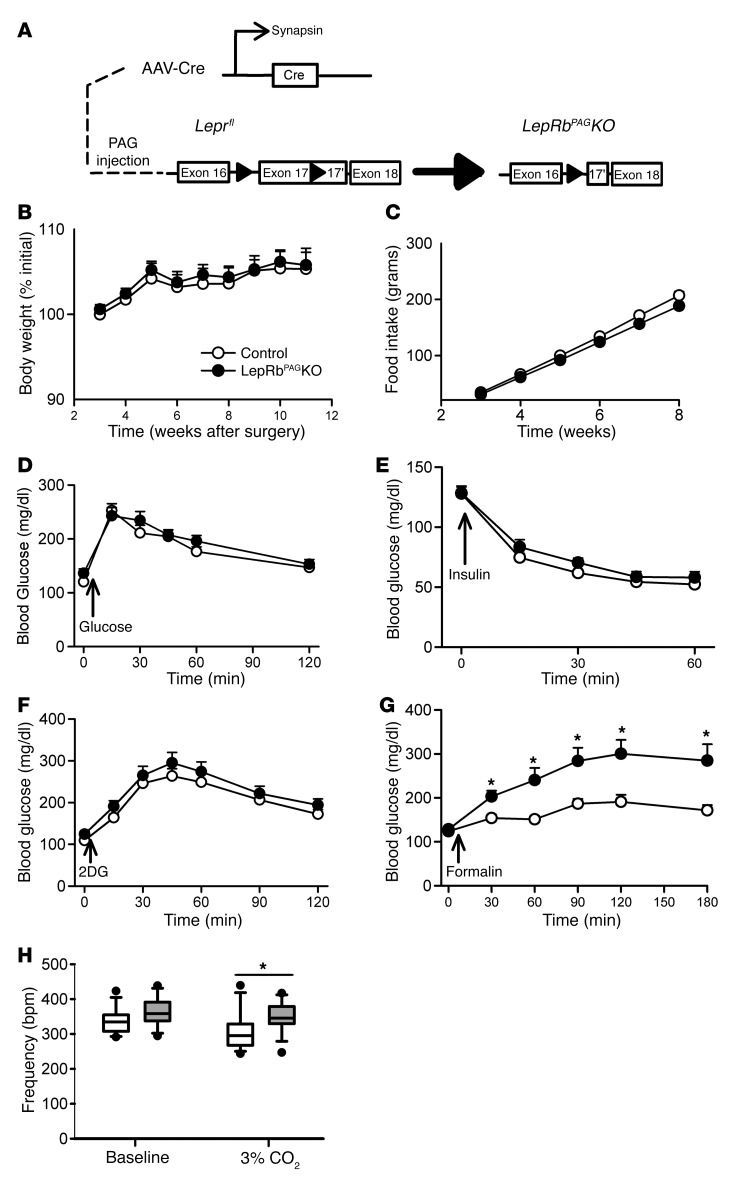
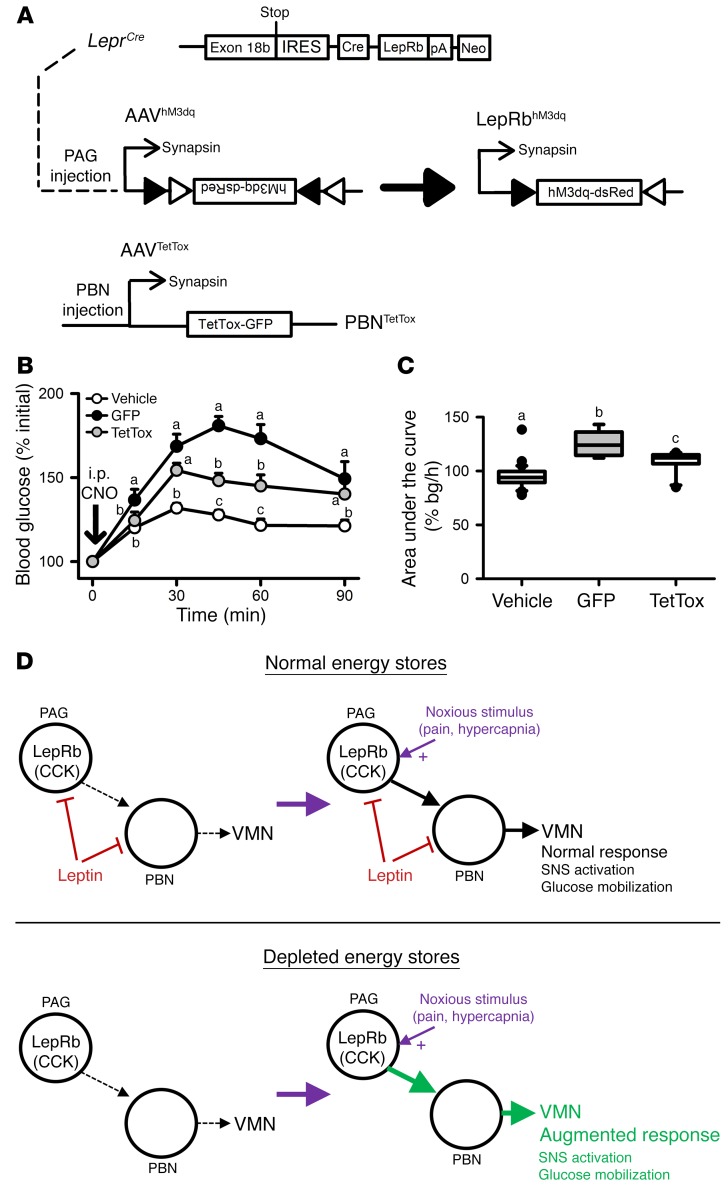
Adipocytes secrete the hormone leptin to signal the sufficiency of energy stores. Reductions in circulating leptin concentrations reflect a negative energy balance, which augments sympathetic nervous system (SNS) activation in response to metabolically demanding emergencies. This process ensures adequate glucose mobilization despite low energy stores. We report that leptin receptor-expressing neurons (LepRb neurons) in the periaqueductal gray (PAG), the largest population of LepRb neurons in the brain stem, mediate this process. Application of noxious stimuli, which often signal the need to mobilize glucose to support an appropriate response, activated PAG LepRb neurons, which project to and activate parabrachial nucleus (PBN) neurons that control SNS activation and glucose mobilization. Furthermore, activating PAG LepRb neurons increased SNS activity and blood glucose concentrations, while ablating LepRb in PAG neurons augmented glucose mobilization in response to noxious stimuli. Thus, decreased leptin action on PAG LepRb neurons augments the autonomic response to noxious stimuli, ensuring sufficient glucose mobilization during periods of acute demand in the face of diminished energy stores.
Conflict of interest statement
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
