

The role of microglial P2X7: modulation of cell death and cytokine release
- PMID: 28716092
- PMCID: PMC5513370
- DOI: 10.1186/s12974-017-0904-8
The role of microglial P2X7: modulation of cell death and cytokine release
Abstract
Background: ATP-gated P2X7 is a non-selective cation channel, which participates in a wide range of cellular functions as well as pathophysiological processes including neuropathic pain, immune response, and neuroinflammation. Despite its abundant expression in microglia, the role of P2X7 in neuroinflammation still remains unclear.
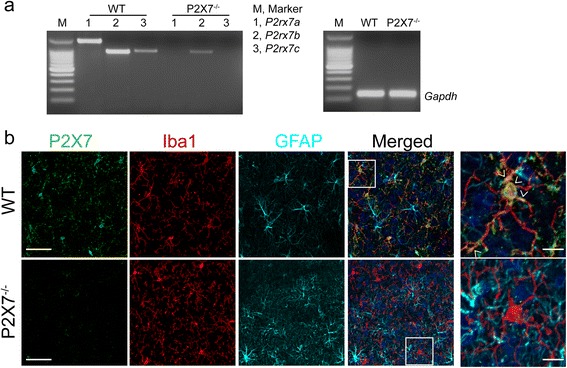
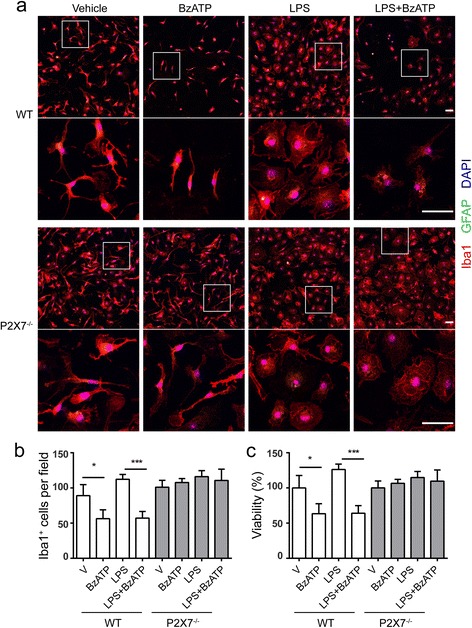
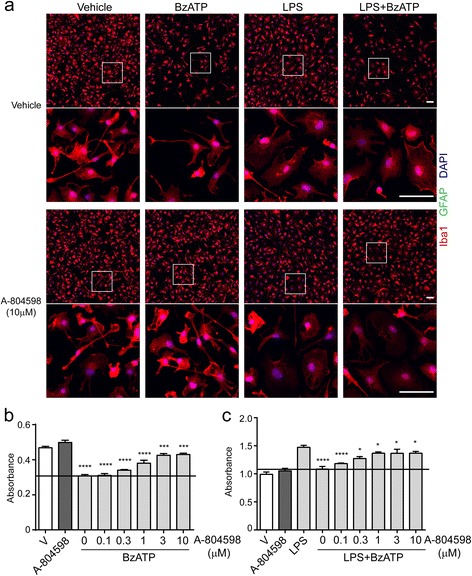
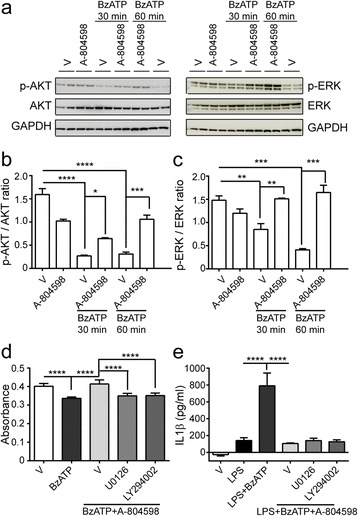
Methods: Primary microglia were isolated from cortices of P0-2 C57BL/6 wild-type or P2X7 knockout (P2X7-/-) mouse pups. Lipopolysaccharide, lipopolysaccharide plus IFNγ, or IL4 plus IL13 were used to polarize microglia to pro-inflammatory or anti-inflammatory states. P2rx7 expression level in resting or activated mouse and human microglia was measured by RNA-sequencing and quantitative real-time PCR. Microglial cell death was measured by cell counting kit-8 and immunocytochemistry, and microglial secretion in wild-type or P2X7-/- microglia was examined by Luminex multiplex assay or ELISA using P2X7 agonist BzATP or P2X7 antagonist A-804598. P2X7 signaling was analyzed by Western blot.
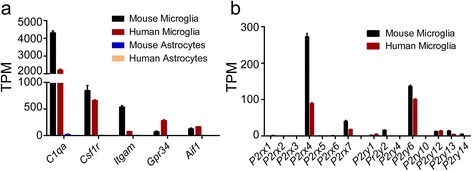
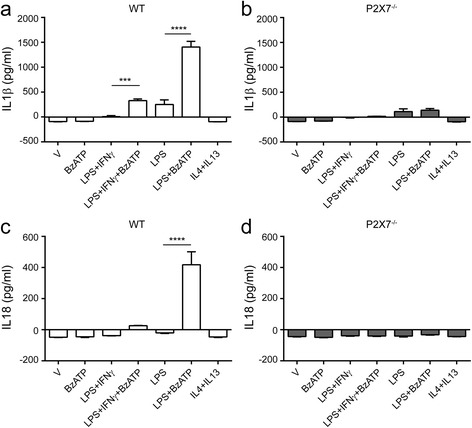
Results: First, we confirmed that P2rx7 is constitutively expressed in mouse and human primary microglia. Moreover, P2rx7 mRNA level was downregulated in mouse microglia under both pro- and anti-inflammatory conditions. Second, P2X7 agonist BzATP caused cell death of mouse microglia, while this effect was suppressed either by P2X7 knockout or by A-804598 under both basal and pro-inflammatory conditions, which suggests the mediating role of P2X7 in BzATP-induced microglial cell death. Third, BzATP-induced release of IL1 family cytokines including IL1α, IL1β, and IL18 was blocked in P2X7-/- microglia or by A-804598 in pro-inflammatory microglia, while the release of other cytokines/chemokines was independent of P2X7 activation. These findings support the specific role of P2X7 in IL1 family cytokine release. Finally, P2X7 activation was discovered to be linked to AKT and ERK pathways, which may be the underlying mechanism of P2X7 functions in microglia.
Conclusions: These results reveal that P2X7 mediates BzATP-induced microglial cell death and specific release of IL1 family cytokines, indicating the important role of P2X7 in neuroinflammation and implying the potential of targeting P2X7 for the treatment of neuroinflammatory disorders.
Keywords: Cell death; Cytokine; Microglia; Neuroinflammation; P2X7.
Figures


References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
