

The characterization of hippocampal theta-driving neurons - a time-delayed mutual information approach
- PMID: 28717183
- PMCID: PMC5514076
- DOI: 10.1038/s41598-017-05527-2
The characterization of hippocampal theta-driving neurons - a time-delayed mutual information approach
Abstract
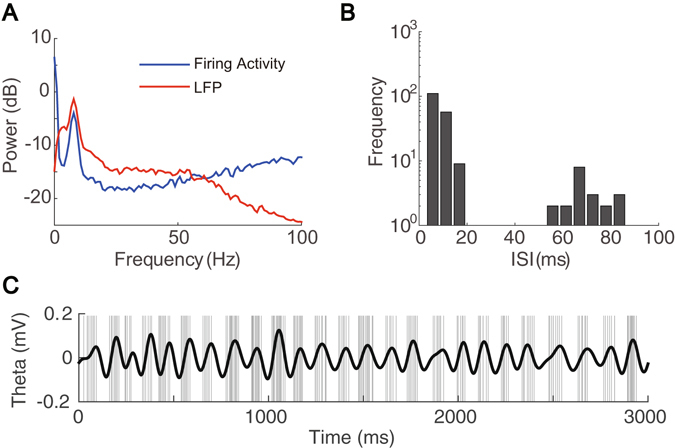
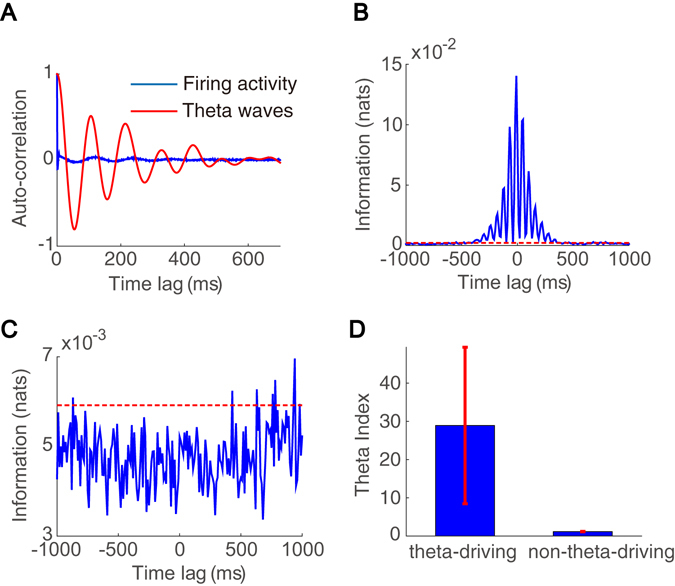
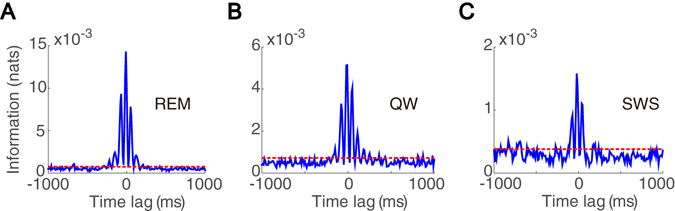
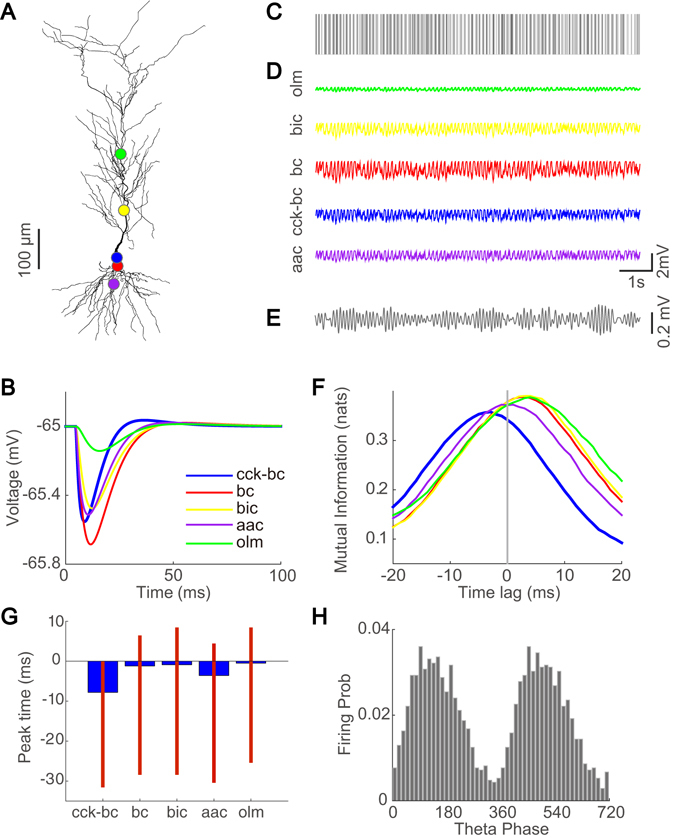
Interneurons are important for computation in the brain, in particular, in the information processing involving the generation of theta oscillations in the hippocampus. Yet the functional role of interneurons in the theta generation remains to be elucidated. Here we use time-delayed mutual information to investigate information flow related to a special class of interneurons-theta-driving neurons in the hippocampal CA1 region of the mouse-to characterize the interactions between theta-driving neurons and theta oscillations. For freely behaving mice, our results show that information flows from the activity of theta-driving neurons to the theta wave, and the firing activity of theta-driving neurons shares a substantial amount of information with the theta wave regardless of behavioral states. Via realistic simulations of a CA1 pyramidal neuron, we further demonstrate that theta-driving neurons possess the characteristics of the cholecystokinin-expressing basket cells (CCK-BC). Our results suggest that it is important to take into account the role of CCK-BC in the generation and information processing of theta oscillations.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
