

Broadly expressed repressors integrate patterning across orthogonal axes in embryos
- PMID: 28720706
- PMCID: PMC5547611
- DOI: 10.1073/pnas.1703001114
Broadly expressed repressors integrate patterning across orthogonal axes in embryos
Abstract
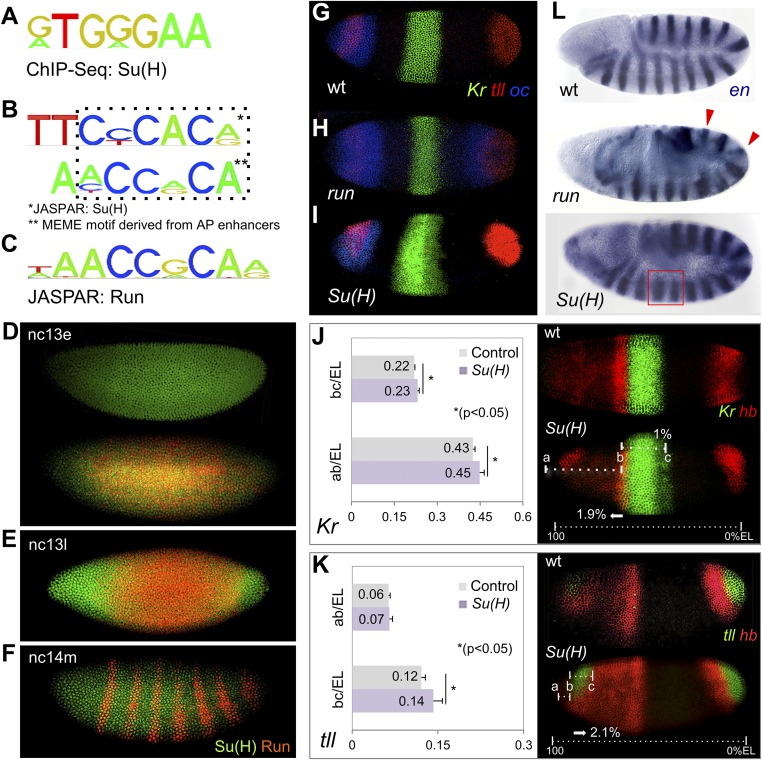
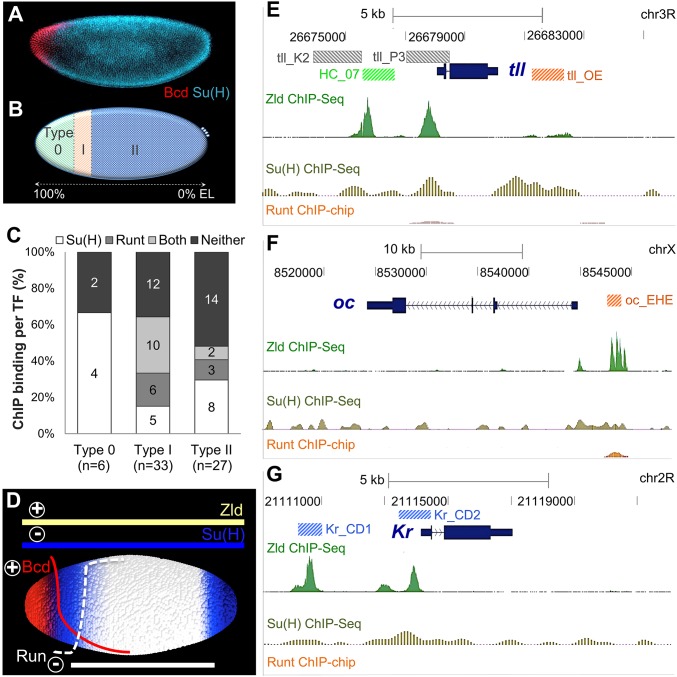
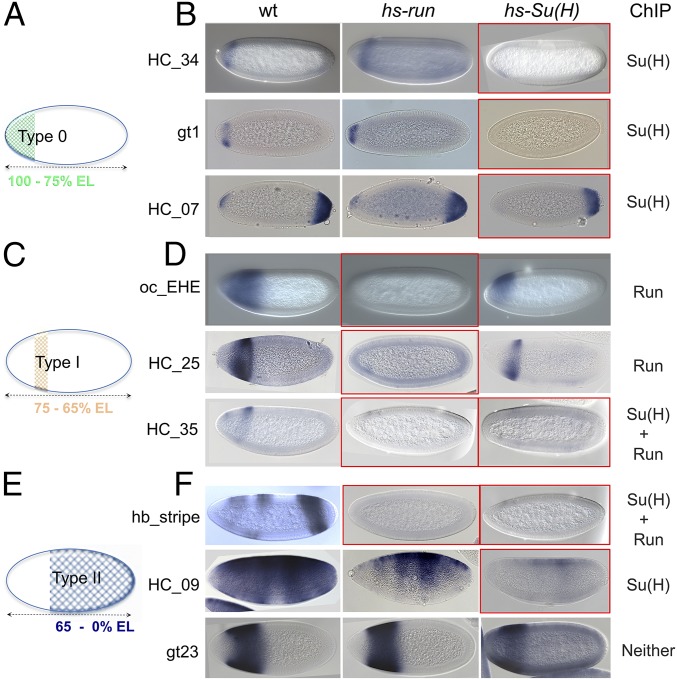
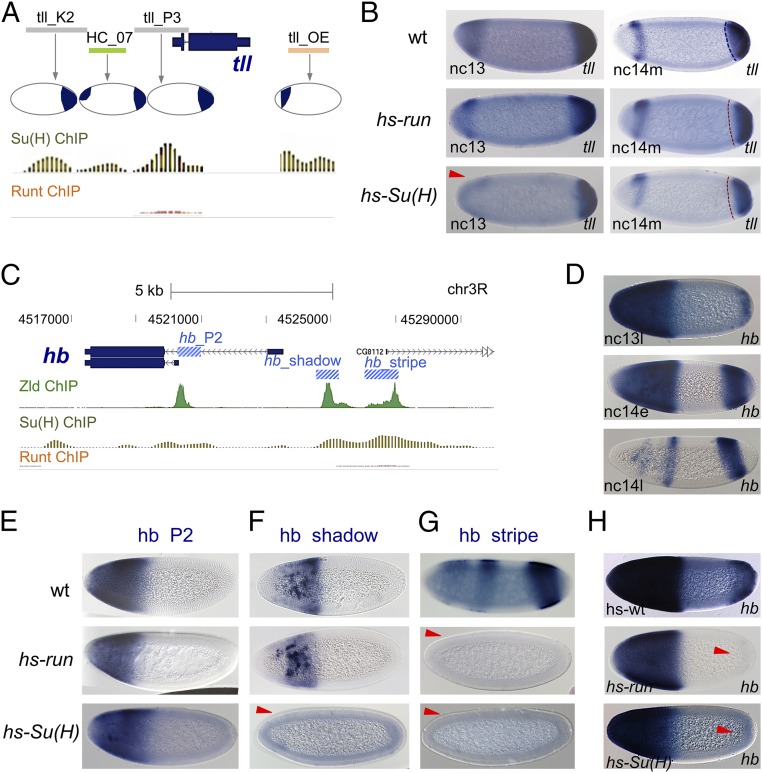
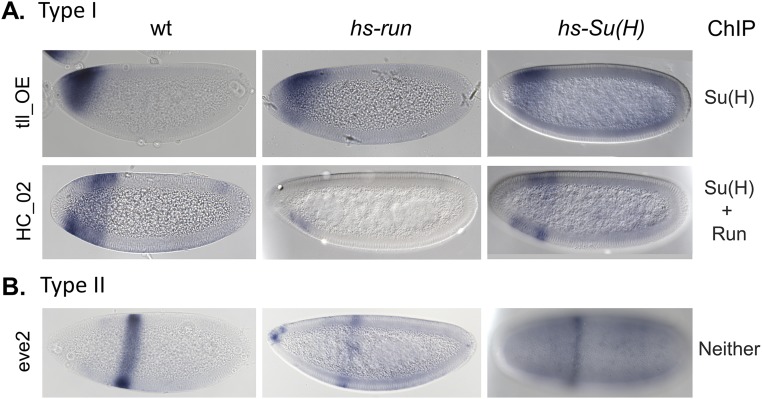
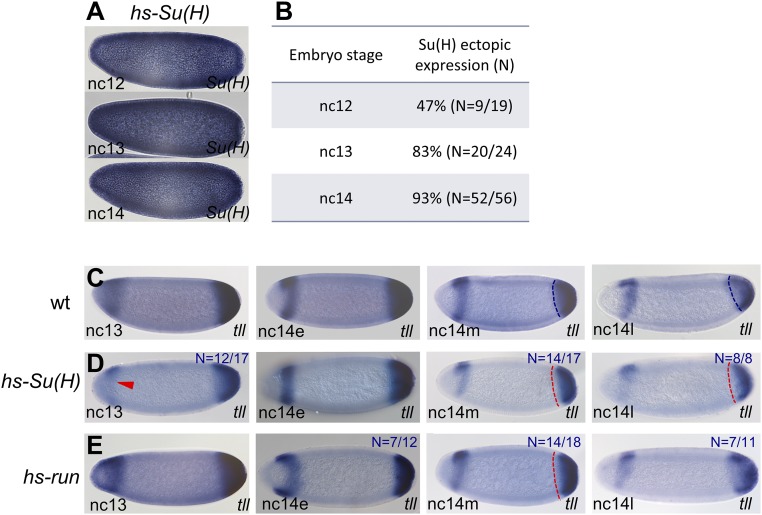
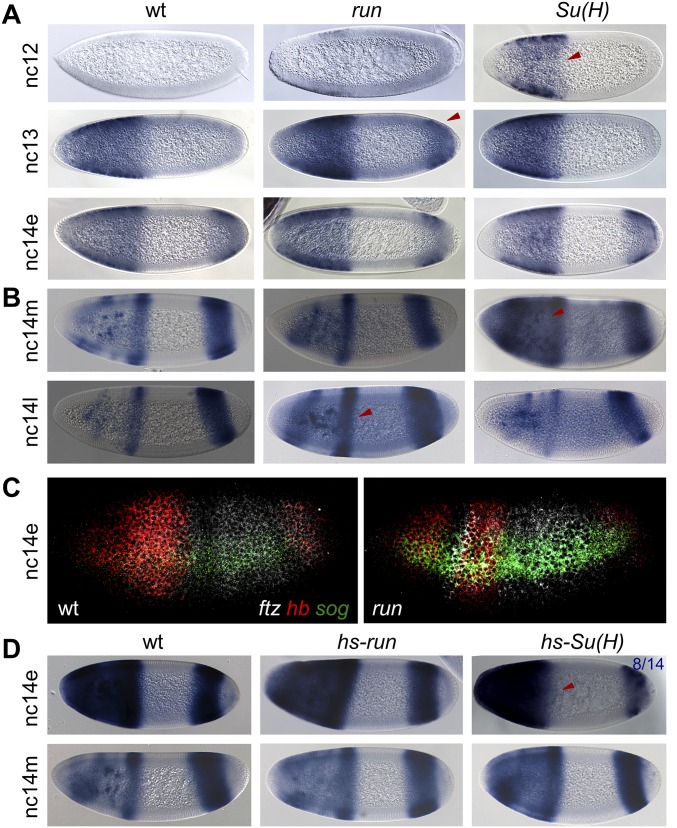
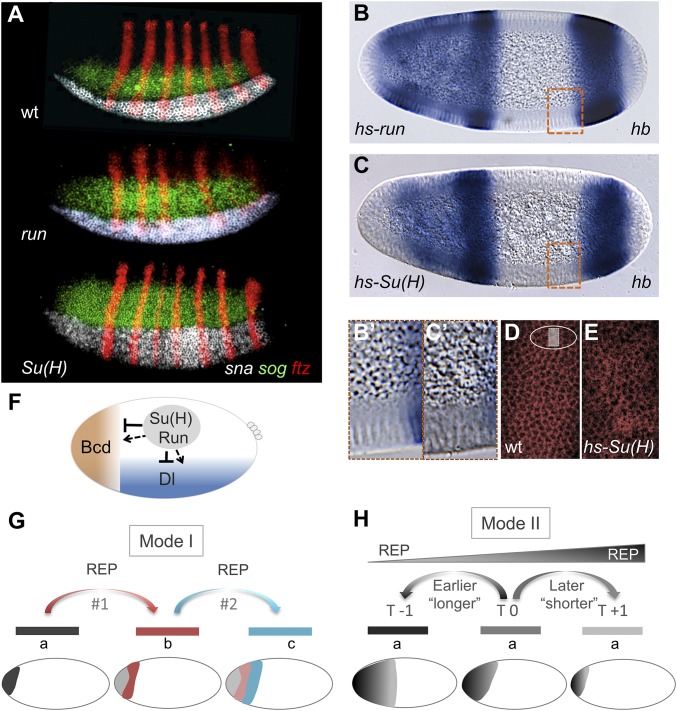
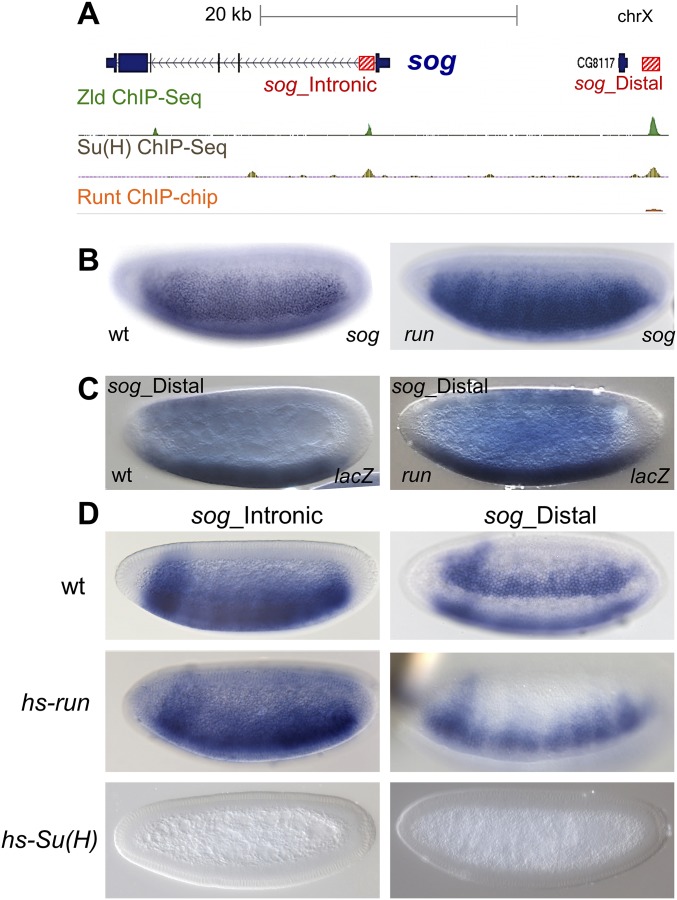
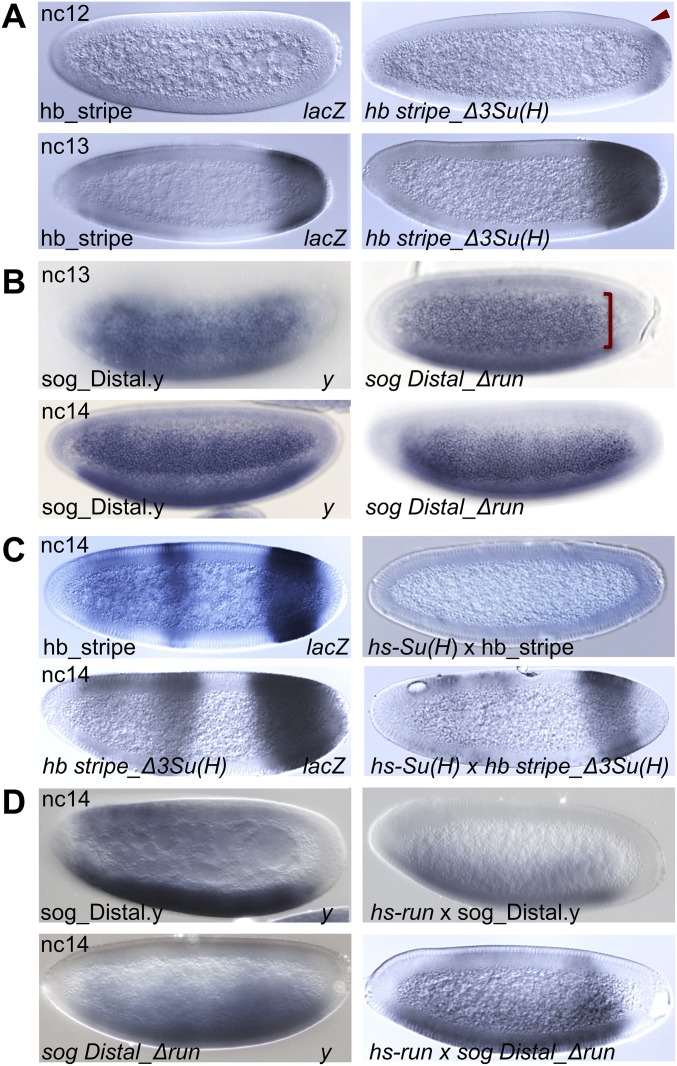
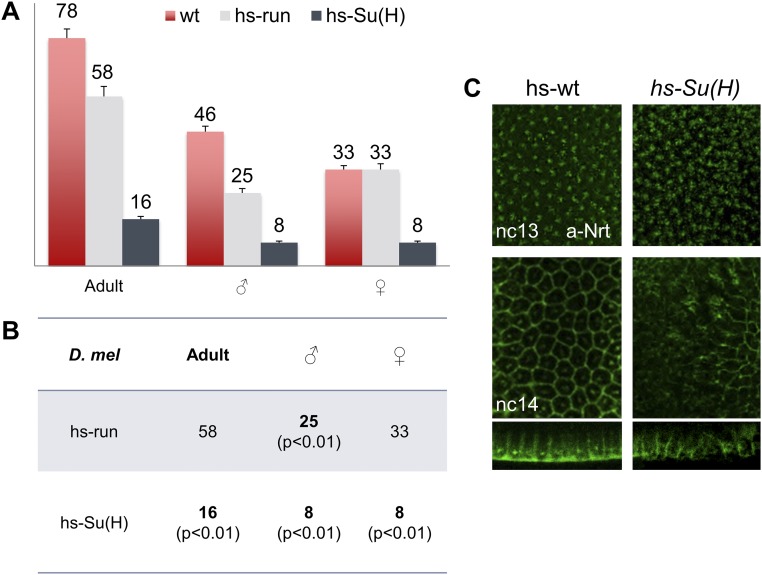
The role of spatially localized repressors in supporting embryonic patterning is well appreciated, but, alternatively, the role ubiquitously expressed repressors play in this process is not well understood. We investigated the function of two broadly expressed repressors, Runt (Run) and Suppressor of Hairless [Su(H)], in patterning the Drosophila embryo. Previous studies have shown that Run and Su(H) regulate gene expression along anterior-posterior (AP) or dorsal-ventral (DV) axes, respectively, by spatially limiting activator action, but here we characterize a different role. Our data show that broadly expressed repressors silence particular enhancers within cis-regulatory systems, blocking their expression throughout the embryo fully but transiently, and, in this manner, regulate spatiotemporal outputs along both axes. Our results suggest that Run and Su(H) regulate the temporal action of enhancers and are not dedicated regulators of one axis but, instead, act coordinately to pattern both axes, AP and DV.
Keywords: Runt; Su(H); embryonic patterning; enhancers; transcriptional repressor.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Rushlow CA, Shvartsman SY. Temporal dynamics, spatial range, and transcriptional interpretation of the Dorsal morphogen gradient. Curr Opin Genet Dev. 2012;22:542–546. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
