

Coronavirus nucleocapsid proteins assemble constitutively in high molecular oligomers
- PMID: 28720894
- PMCID: PMC5515880
- DOI: 10.1038/s41598-017-06062-w
Coronavirus nucleocapsid proteins assemble constitutively in high molecular oligomers
Abstract
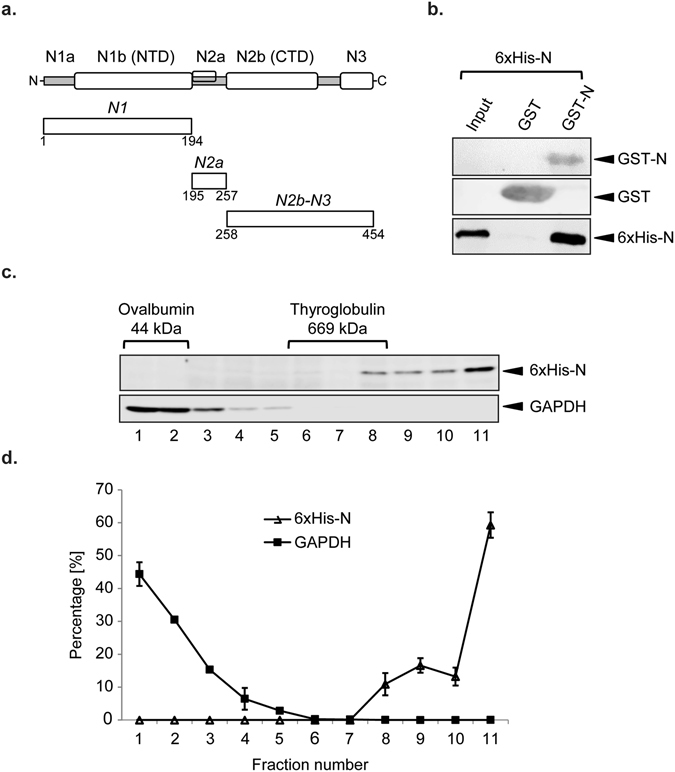
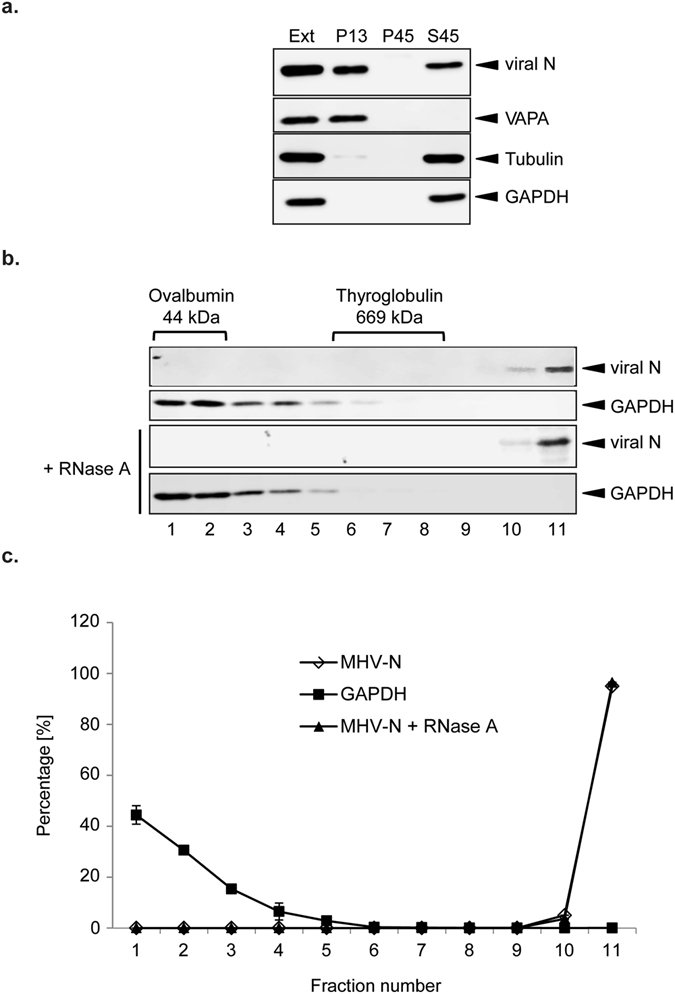
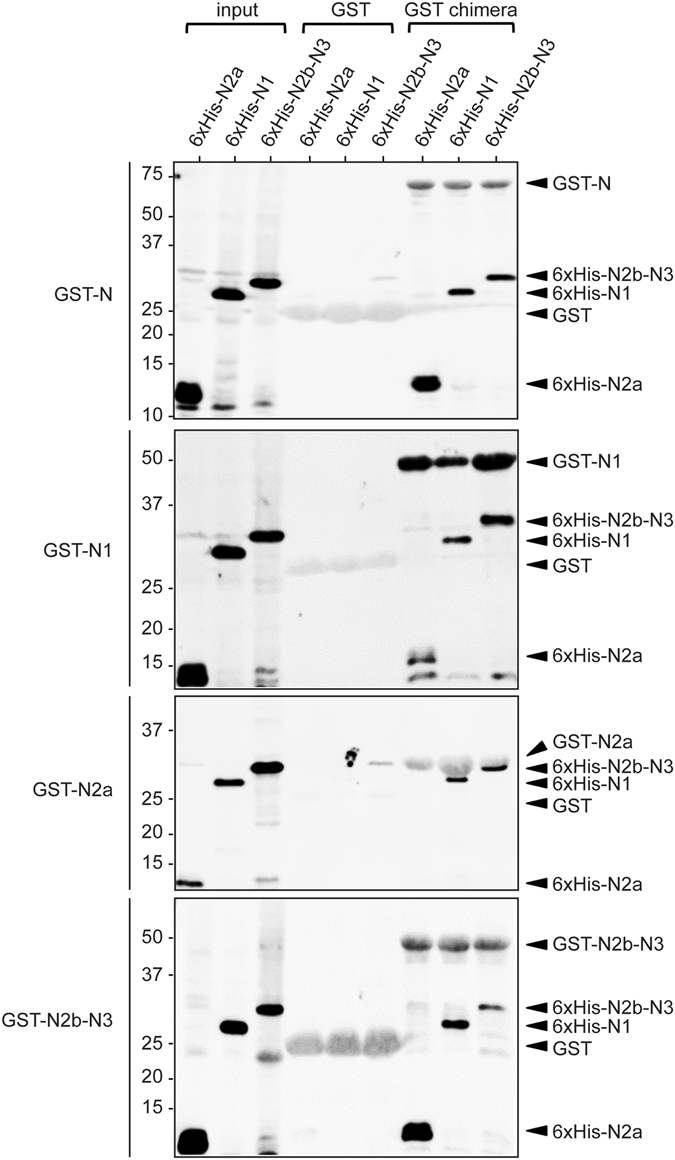
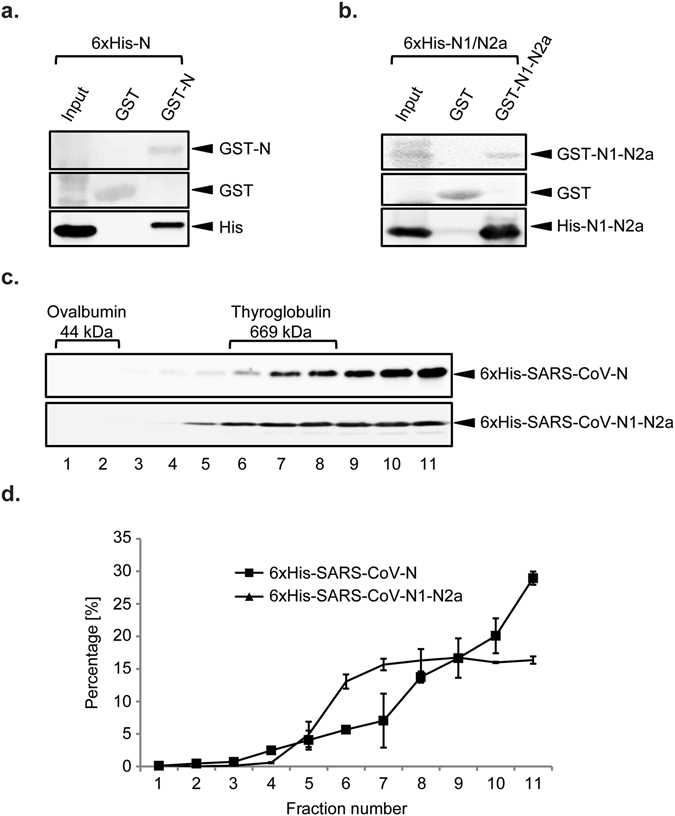
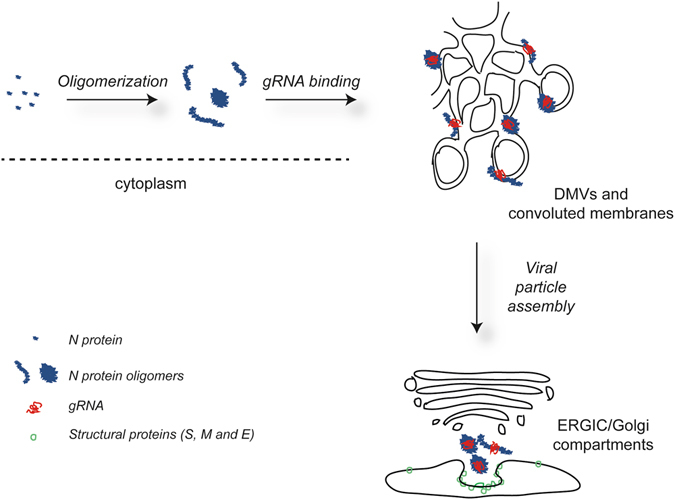
Coronaviruses (CoV) are enveloped viruses and rely on their nucleocapsid N protein to incorporate the positive-stranded genomic RNA into the virions. CoV N proteins form oligomers but the mechanism and relevance underlying their multimerization remain to be fully understood. Using in vitro pull-down experiments and density glycerol gradients, we found that at least 3 regions distributed over its entire length mediate the self-interaction of mouse hepatitis virus (MHV) and severe acute respiratory syndrome coronavirus (SARS-CoV) N protein. The fact that these regions can bind reciprocally between themselves provides a possible molecular basis for N protein oligomerization. Interestingly, cytoplasmic N molecules of MHV-infected cells constitutively assemble into oligomers through a process that does not require binding to genomic RNA. Based on our data, we propose a model where constitutive N protein oligomerization allows the optimal loading of the genomic viral RNA into a ribonucleoprotein complex via the presentation of multiple viral RNA binding motifs.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
