

Current Perspectives for the use of Gonane Progesteronergic Drugs in the Treatment of Central Hypoventilation Syndromes
- PMID: 28721821
- PMCID: PMC6295933
- DOI: 10.2174/1570159X15666170719104605
Current Perspectives for the use of Gonane Progesteronergic Drugs in the Treatment of Central Hypoventilation Syndromes
Abstract
Background: Central alveolar hypoventilation syndromes (CHS) encompass neurorespiratory diseases resulting from congenital or acquired neurological disorders. Hypercapnia, acidosis, and hypoxemia resulting from CHS negatively affect physiological functions and can be lifethreatening. To date, the absence of pharmacological treatment implies that the patients must receive assisted ventilation throughout their lives.
Objective: To highlight the relevance of determining conditions in which using gonane synthetic progestins could be of potential clinical interest for the treatment of CHS.
Methods: The mechanisms by which gonanes modulate the respiratory drive were put into the context of those established for natural progesterone and other synthetic progestins.
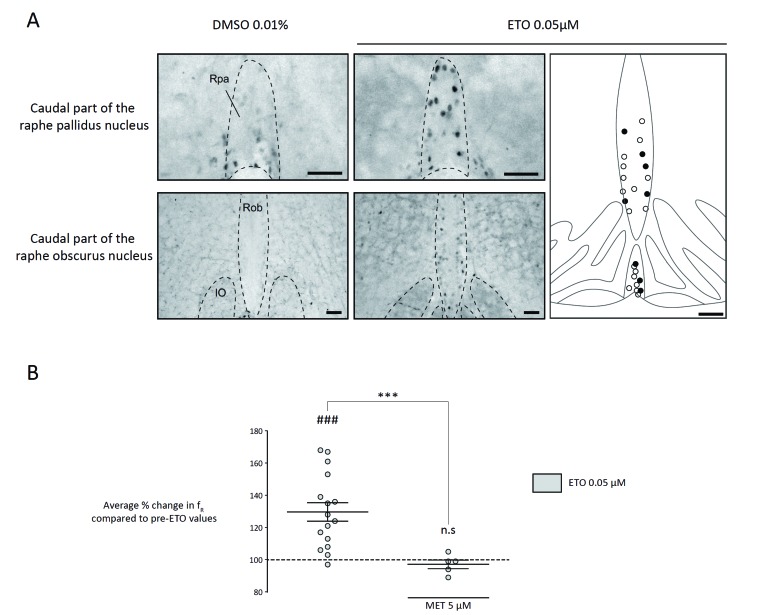
Results: The clinical benefits of synthetic progestins to treat respiratory diseases are mixed with either positive outcomes or no improvement. A benefit for CHS patients has only recently been proposed. We incidentally observed restoration of CO2 chemosensitivity, the functional deficit of this disease, in two adult CHS women by desogestrel, a gonane progestin, used for contraception. This effect was not observed by another group, studying a single patient. These contradictory findings are probably due to the complex nature of the action of desogestrel on breathing and led us to carry out mechanistic studies in rodents. Our results show that desogestrel influences the respiratory command by modulating the GABAA and NMDA signaling in the respiratory network, medullary serotoninergic systems, and supramedullary areas.
Conclusion: Gonanes show promise for improving ventilation of CHS patients, although the conditions of their use need to be better understood.
Keywords: Central congenital hypoventilation syndrome; Ondine’s curse syndrome; desogestrel; etonogestrel; gonane; progesterone; respiratory control..
Copyright© Bentham Science Publishers; For any queries, please email at epub@benthamscience.org.
Figures




Similar articles
-
Serotonin and the ventilatory effects of etonogestrel, a gonane progestin, in a murine model of congenital central hypoventilation syndrome.Front Endocrinol (Lausanne). 2023 Feb 21;14:1077798. doi: 10.3389/fendo.2023.1077798. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 36896185 Free PMC article.
-
[Alone or combined with serotoninergic agents, the progestins of the gonane family may represent a therapeutic alternative for Ondine's curse syndrome].Rev Mal Respir. 2022 Feb;39(2):95-99. doi: 10.1016/j.rmr.2022.01.008. Epub 2022 Feb 11. Rev Mal Respir. 2022. PMID: 35165014 French.
-
Desogestrel enhances ventilation in ondine patients: Animal data involving serotoninergic systems.Neuropharmacology. 2016 Aug;107:339-350. doi: 10.1016/j.neuropharm.2016.03.041. Epub 2016 Apr 1. Neuropharmacology. 2016. PMID: 27040794
-
Congenital central hypoventilation syndrome and desogestrel: a call for caution: addendum to "C. Straus, H. Trang, M.H. Becquemin, P. Touraine, T. Similowski, Chemosensitivity recovery in Ondine's curse syndrome under treatment with desogestrel" [Respir. Physiol. Neurobiol. 171 (2010) 171-174].Respir Physiol Neurobiol. 2011 Sep 15;178(2):357-8. doi: 10.1016/j.resp.2011.07.007. Epub 2011 Jul 23. Respir Physiol Neurobiol. 2011. PMID: 21801857 Review.
-
Comparative pharmacology of newer progestogens.Drugs. 1996 Feb;51(2):188-215. doi: 10.2165/00003495-199651020-00002. Drugs. 1996. PMID: 8808163 Review.
Cited by
-
Serotonin and the ventilatory effects of etonogestrel, a gonane progestin, in a murine model of congenital central hypoventilation syndrome.Front Endocrinol (Lausanne). 2023 Feb 21;14:1077798. doi: 10.3389/fendo.2023.1077798. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 36896185 Free PMC article.
-
Evaluation of the effectiveness of medroxyprogesterone on blood gases and short-term hospital outcomes in patients with chronic obstructive pulmonary disease treating with noninvasive ventilation: A randomized clinical trial.J Res Med Sci. 2024 Nov 28;29:72. doi: 10.4103/jrms.jrms_198_24. eCollection 2024. J Res Med Sci. 2024. PMID: 39764220 Free PMC article.
-
Etonogestrel Administration Reduces the Expression of PHOX2B and Its Target Genes in the Solitary Tract Nucleus.Int J Mol Sci. 2022 Apr 27;23(9):4816. doi: 10.3390/ijms23094816. Int J Mol Sci. 2022. PMID: 35563209 Free PMC article.
-
In Transgenic Erythropoietin Deficient Mice, an Increase in Respiratory Response to Hypercapnia Parallels Abnormal Distribution of CO2/H+-Activated Cells in the Medulla Oblongata.Front Physiol. 2022 Apr 19;13:850418. doi: 10.3389/fphys.2022.850418. eCollection 2022. Front Physiol. 2022. PMID: 35514353 Free PMC article.
-
Research Advances on Therapeutic Approaches to Congenital Central Hypoventilation Syndrome (CCHS).Front Neurosci. 2021 Jan 12;14:615666. doi: 10.3389/fnins.2020.615666. eCollection 2020. Front Neurosci. 2021. PMID: 33510615 Free PMC article. Review.
References
-
- Feldman J.L., Del Negro C.A., Gray P.A. Understanding the rhythm of breathing: so near, yet so far. Annu. Rev. Physiol. 2013;75:423–452. [http://dx.doi.org/10.1146/annurev-physiol-040510-130049]. [PMID: 23121137]. - PMC - PubMed
-
- Feldman J.L., Kam K. Facing the challenge of mammalian neural microcircuits: taking a few breaths may help. J. Physiol. 2015;593(1):3–23. [http://dx.doi.org/10.1113/jphysiol.2014.277632]. [PMID: 25556783]. - PMC - PubMed
-
- Richter D.W., Smith J.C. Respiratory rhythm generation in vivo. Physiology (Bethesda) 2014;29(1):58–71. [http://dx.doi.org/10. 1152/physiol.00035.2013]. [PMID: 24382872]. - PMC - PubMed
-
- Smith J.C., Ellenberger H.H., Ballanyi K., Richter D.W.
- Feldman J.L. Pre-Bötzinger complex: a brainstem region that may generate respiratory rhythm in mammals. Science. 1991;254(5032):726–729. [http://dx.doi.org/10.1126/science.1683005]. [PMID: 1683005]. - PMC - PubMed
-
- Onimaru H., Homma I. A novel functional neuron group for respiratory rhythm generation in the ventral medulla. J. Neurosci. 2003;23(4):1478–1486. [http://dx.doi.org/10.1523/JNEUROSCI.23-04-01478.2003]. [PMID: 12598636]. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources