

Termination factor Rho: From the control of pervasive transcription to cell fate determination in Bacillus subtilis
- PMID: 28723971
- PMCID: PMC5540618
- DOI: 10.1371/journal.pgen.1006909
Termination factor Rho: From the control of pervasive transcription to cell fate determination in Bacillus subtilis
Abstract
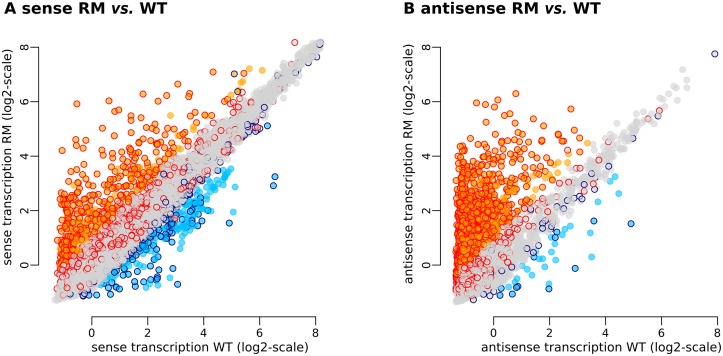
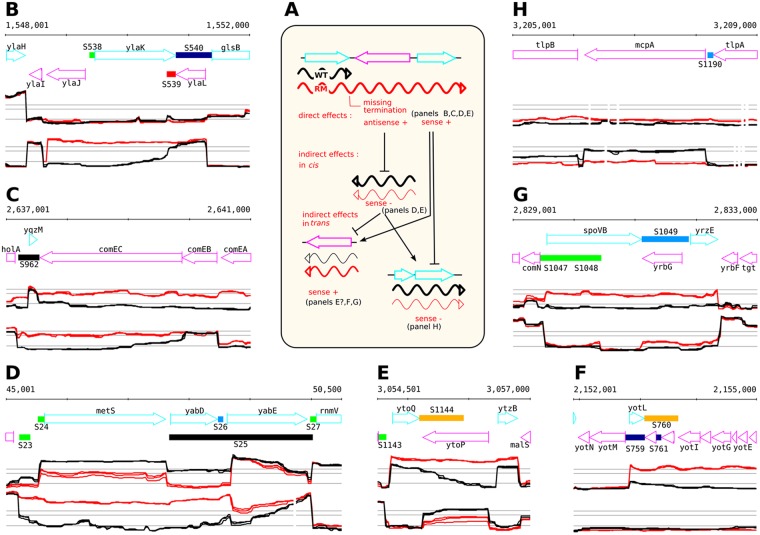
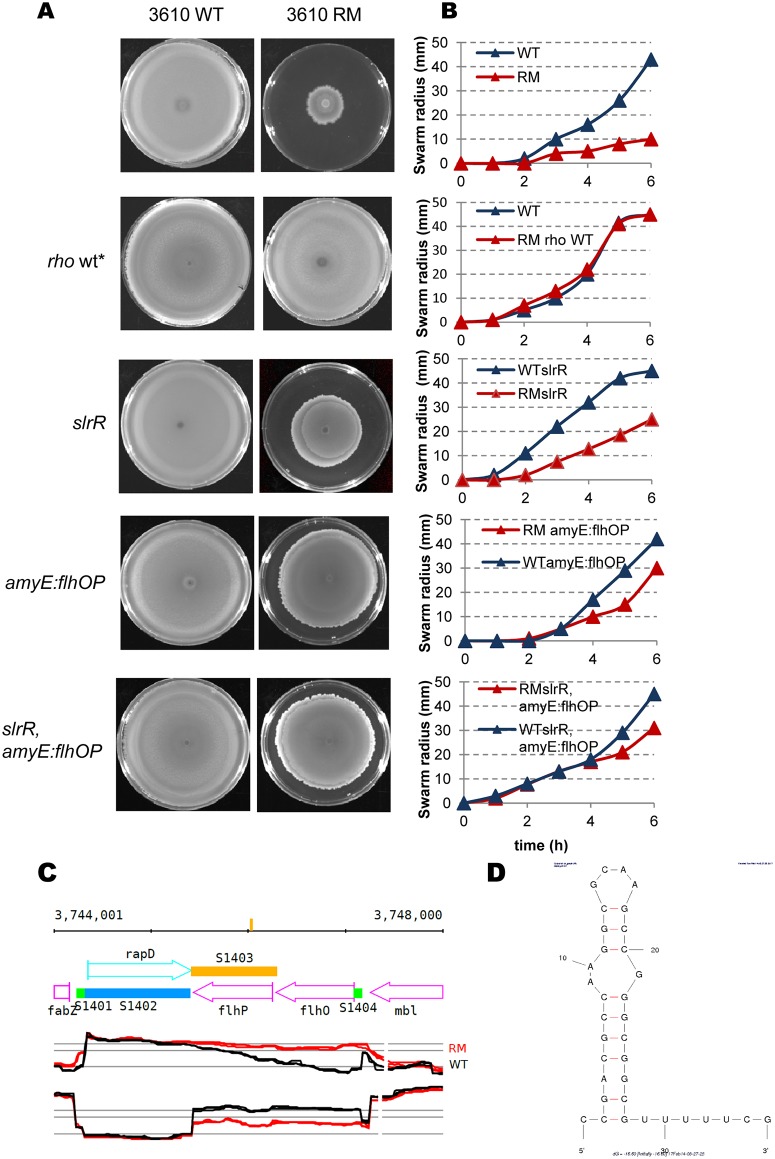
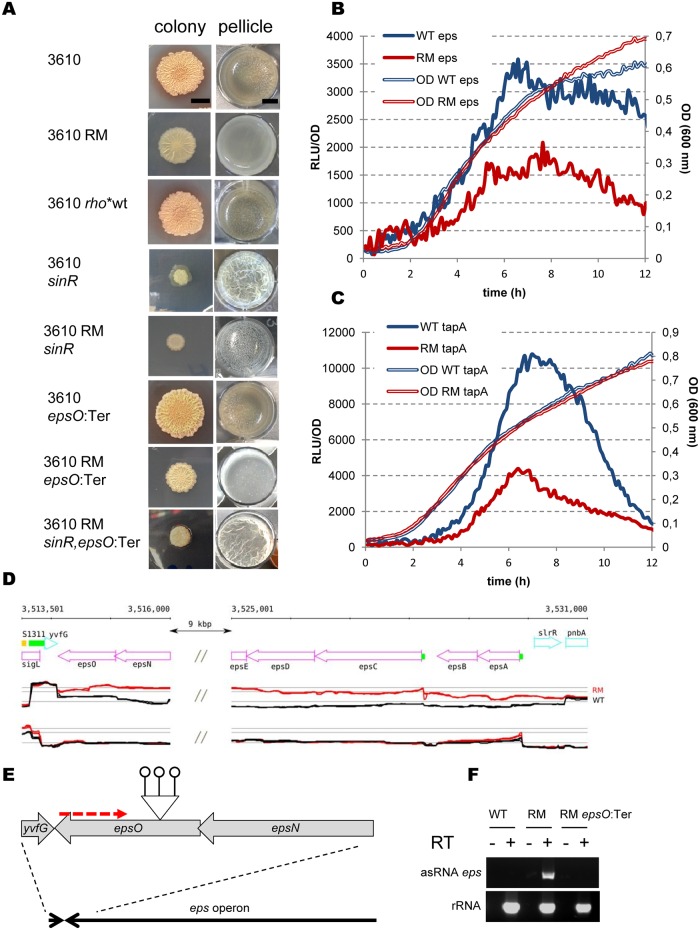
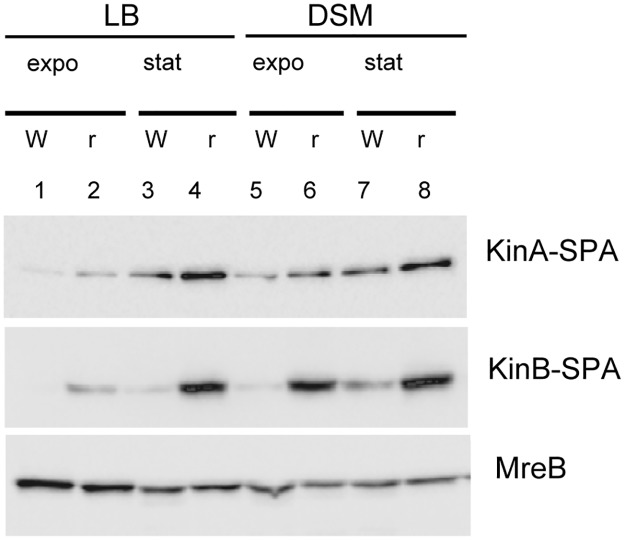
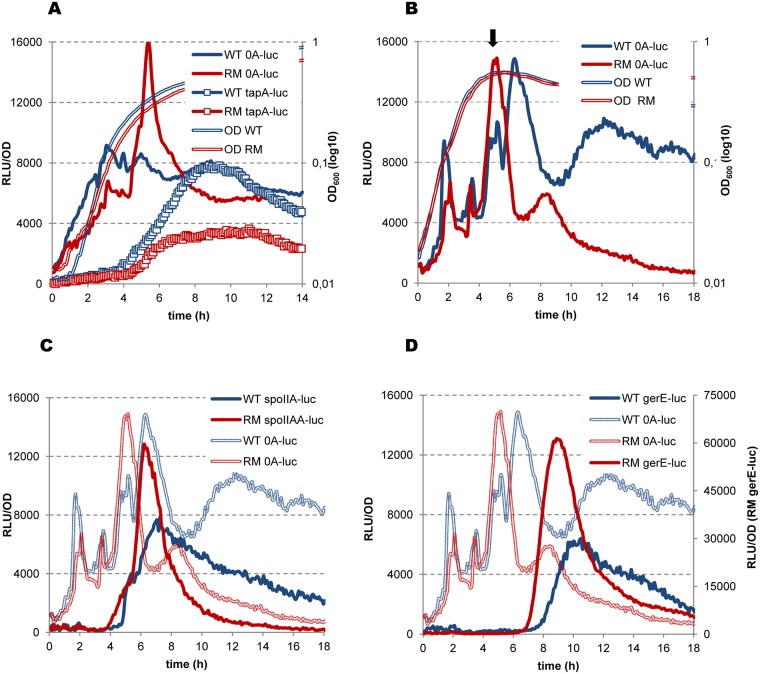
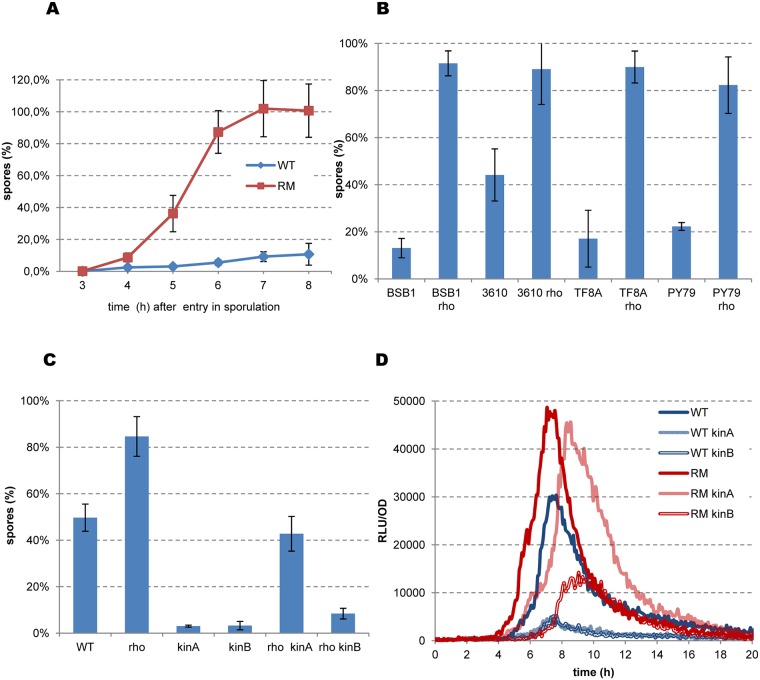
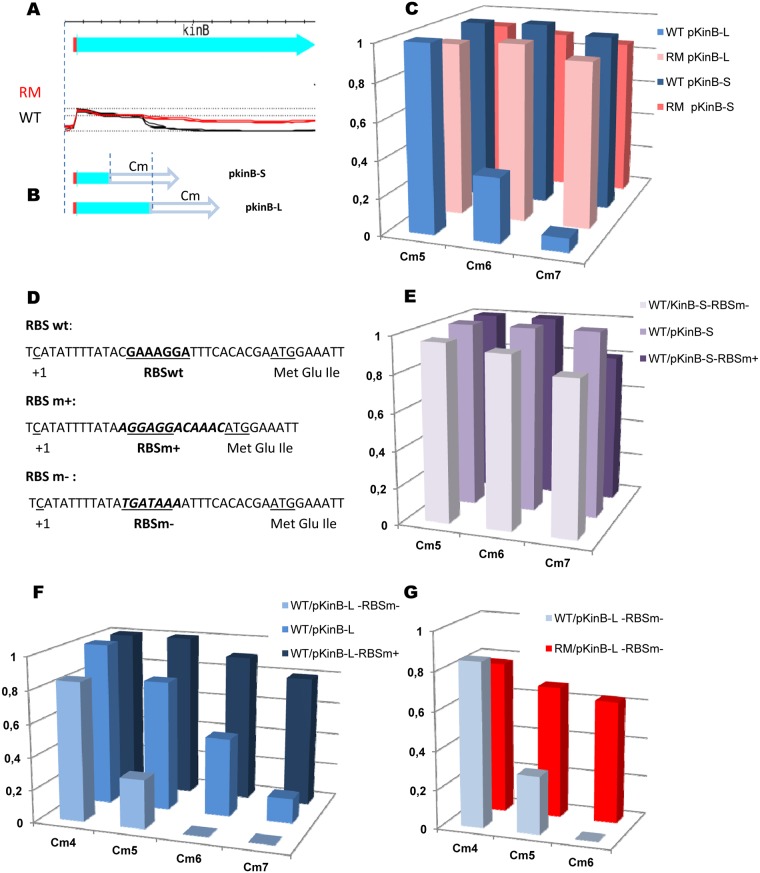
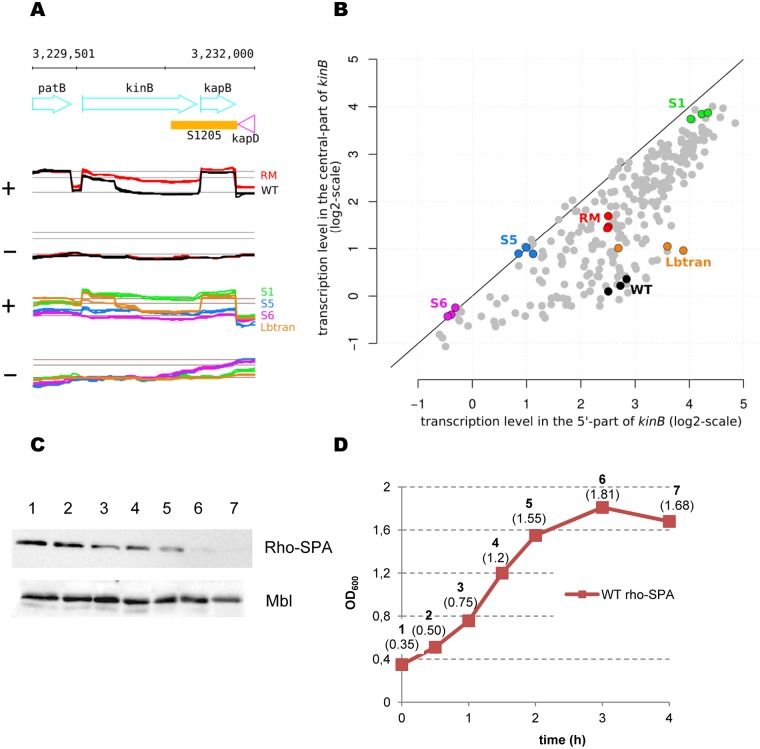
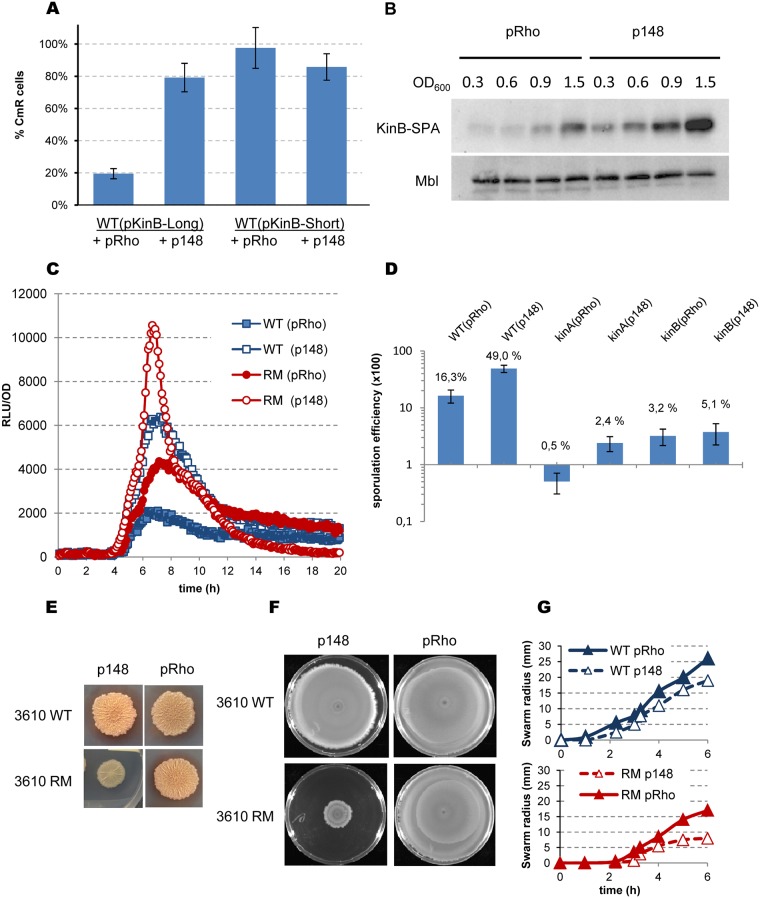
In eukaryotes, RNA species originating from pervasive transcription are regulators of various cellular processes, from the expression of individual genes to the control of cellular development and oncogenesis. In prokaryotes, the function of pervasive transcription and its output on cell physiology is still unknown. Most bacteria possess termination factor Rho, which represses pervasive, mostly antisense, transcription. Here, we investigate the biological significance of Rho-controlled transcription in the Gram-positive model bacterium Bacillus subtilis. Rho inactivation strongly affected gene expression in B. subtilis, as assessed by transcriptome and proteome analysis of a rho-null mutant during exponential growth in rich medium. Subsequent physiological analyses demonstrated that a considerable part of Rho-controlled transcription is connected to balanced regulation of three mutually exclusive differentiation programs: cell motility, biofilm formation, and sporulation. In the absence of Rho, several up-regulated sense and antisense transcripts affect key structural and regulatory elements of these differentiation programs, thereby suppressing motility and biofilm formation and stimulating sporulation. We dissected how Rho is involved in the activity of the cell fate decision-making network, centered on the master regulator Spo0A. We also revealed a novel regulatory mechanism of Spo0A activation through Rho-dependent intragenic transcription termination of the protein kinase kinB gene. Altogether, our findings indicate that distinct Rho-controlled transcripts are functional and constitute a previously unknown built-in module for the control of cell differentiation in B. subtilis. In a broader context, our results highlight the recruitment of the termination factor Rho, for which the conserved biological role is probably to repress pervasive transcription, in highly integrated, bacterium-specific, regulatory networks.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Libri D. Sleeping Beauty and the Beast (of pervasive transcription). RNA. 2015; 21: 678–679. doi: 10.1261/rna.050948.115 - DOI - PMC - PubMed
-
- Kapranov P, Willingham AT, Gingeras TR. Genome-wide transcription and the implications for genomic organization. Nat Rev Genet. 2007; 8: 413–423. doi: 10.1038/nrg2083 - DOI - PubMed
-
- Berretta J, Morillon A. Pervasive transcription constitutes a new level of eukaryotic genome regulation. EMBO Rep. 2009; 10: 973–982. doi: 10.1038/embor.2009.181 - DOI - PMC - PubMed
-
- Canny SP, Reese TA, Johnson LS, Zhang X, Kambal A, Duan E, et al. Pervasive transcription of a herpesvirus genome generates functionally important RNAs. MBio. 2014; 5: e01033–13. doi: 10.1128/mBio.01033-13 - DOI - PMC - PubMed
-
- Lybecker M, Bilusic I, & Raghavan R. Pervasive transcription: detecting functional RNAs in bacteria. Transcription. 2014; 5: e944039 doi: 10.4161/21541272.2014.944039 - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
