

Nucleocytoplasmic Shuttling of Histone Deacetylase 9 Controls Activity-Dependent Thalamocortical Axon Branching
- PMID: 28729685
- PMCID: PMC5519695
- DOI: 10.1038/s41598-017-06243-7
Nucleocytoplasmic Shuttling of Histone Deacetylase 9 Controls Activity-Dependent Thalamocortical Axon Branching
Abstract
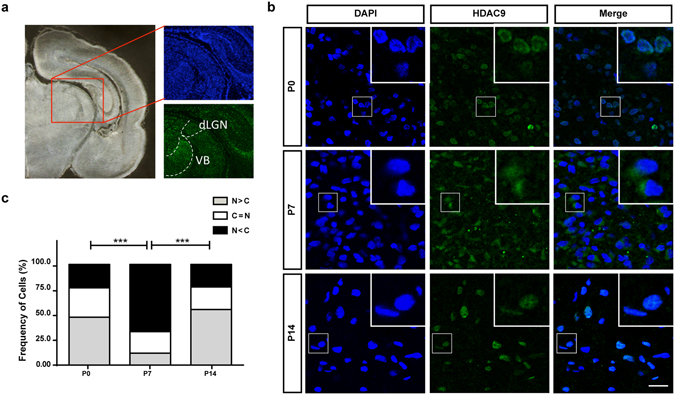
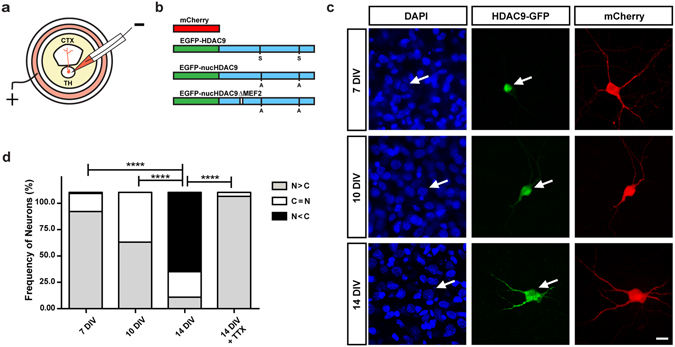
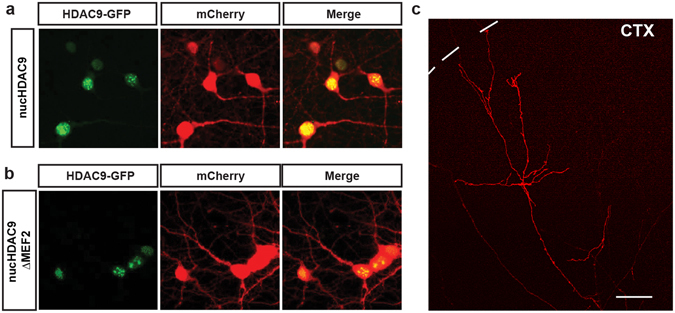
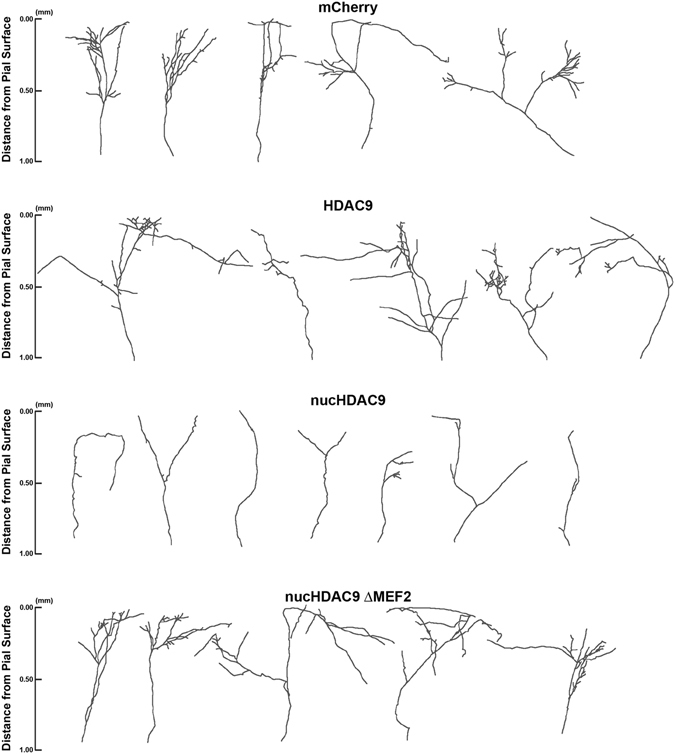
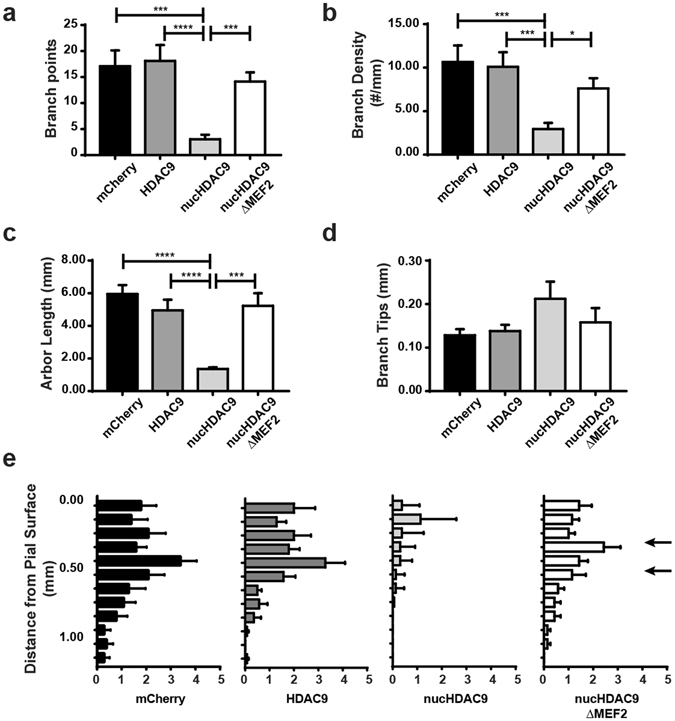
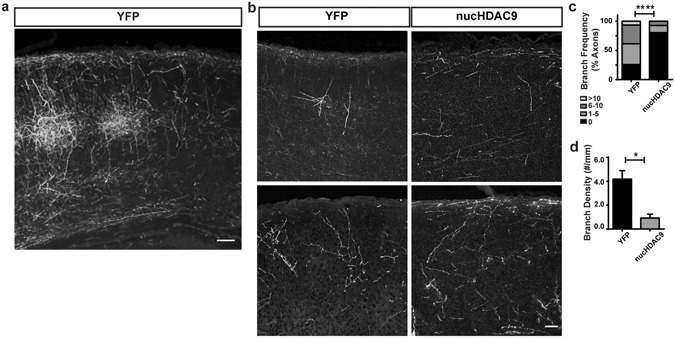
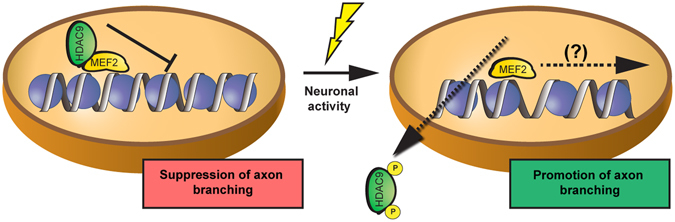
During development, thalamocortical (TC) axons form branches in an activity-dependent fashion. Here we investigated how neuronal activity is converted to molecular signals, focusing on an epigenetic mechanism involving histone deacetylases (HDACs). Immunohistochemistry demonstrated that HDAC9 was translocated from the nucleus to the cytoplasm of thalamic cells during the first postnatal week in rats. In organotypic co-cultures of the thalamus and cortex, fluorescent protein-tagged HDAC9 also exhibited nuclueocytoplasmic translocation in thalamic cells during culturing, which was reversed by tetrodotoxin treatment. Transfection with a mutant HDAC9 that interferes with the translocation markedly decreased TC axon branching in the culture. Similarly, TC axon branching was significantly decreased by the mutant HDAC9 gene transfer in vivo. However, axonal branching was restored by disrupting the interaction between HDAC9 and myocyte-specific enhancer factor 2 (MEF2). Taken together, the present results demonstrate that the nucleocytoplasmic translocation of HDAC9 plays a critical role in activity-dependent TC axon branching by affecting transcriptional regulation and downstream signaling pathways.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
