

Sexually divergent induction of microglial-associated neuroinflammation with hippocampal aging
- PMID: 28732515
- PMCID: PMC5521082
- DOI: 10.1186/s12974-017-0920-8
Sexually divergent induction of microglial-associated neuroinflammation with hippocampal aging
Abstract
Background: The necessity of including both males and females in molecular neuroscience research is now well understood. However, there is relatively limited basic biological data on brain sex differences across the lifespan despite the differences in age-related neurological dysfunction and disease between males and females.
Methods: Whole genome gene expression of young (3 months), adult (12 months), and old (24 months) male and female C57BL6 mice hippocampus was analyzed. Subsequent bioinformatic analyses and confirmations of age-related changes and sex differences in hippocampal gene and protein expression were performed.
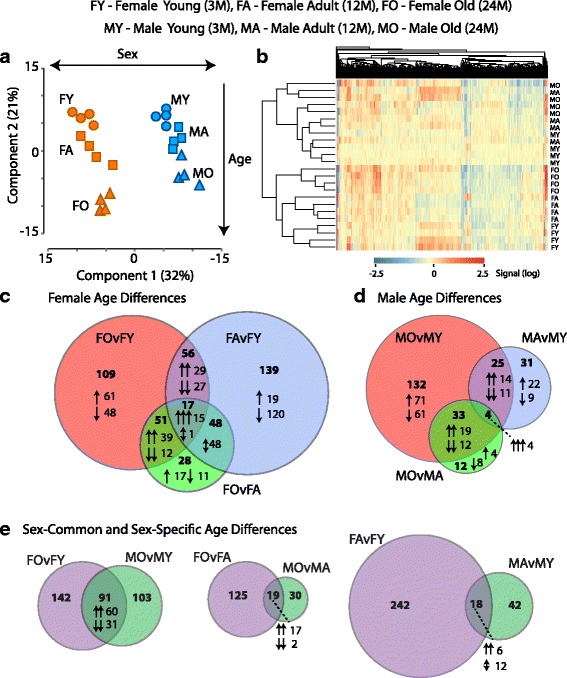
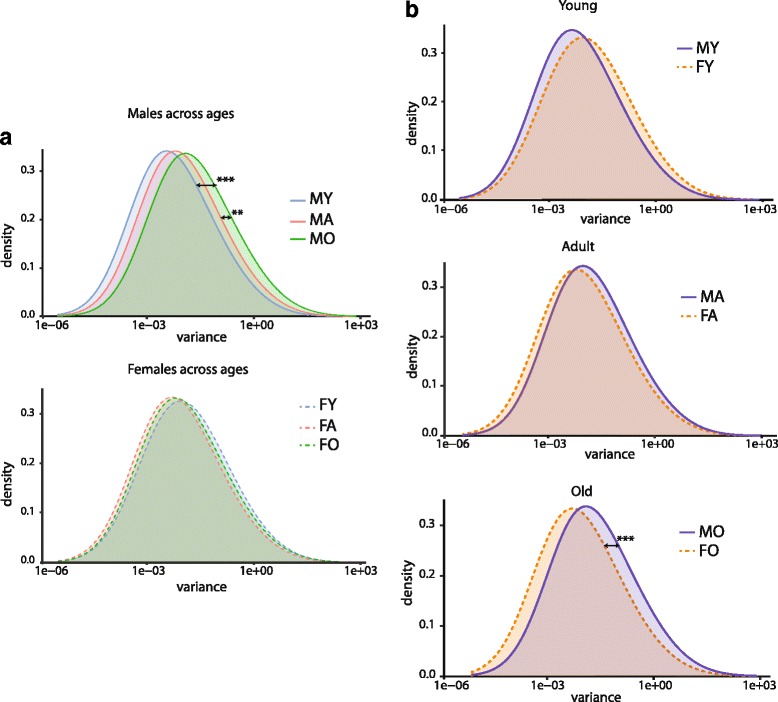
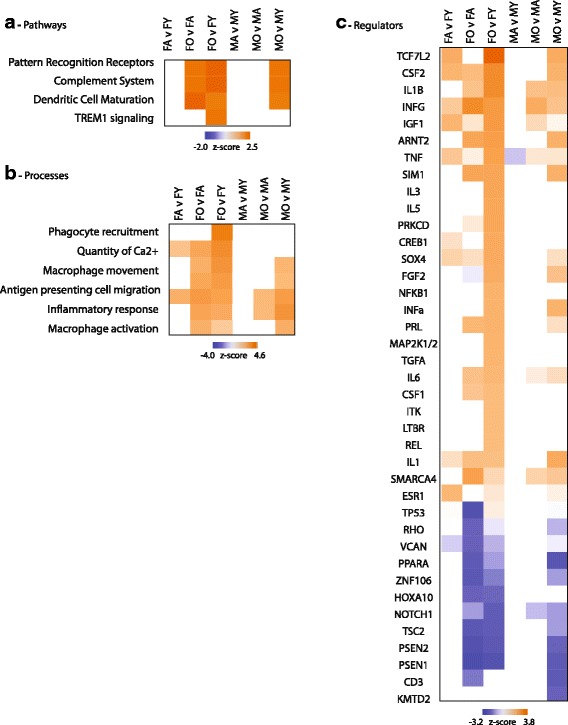
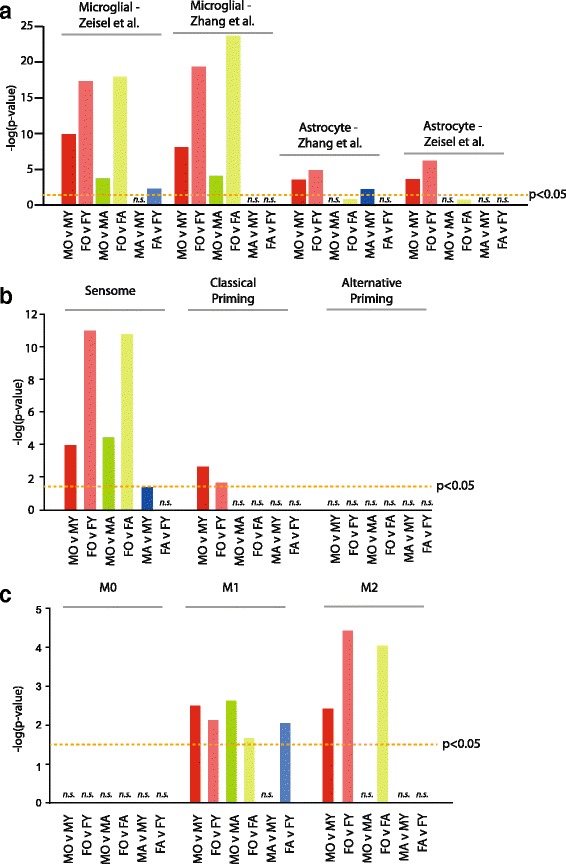
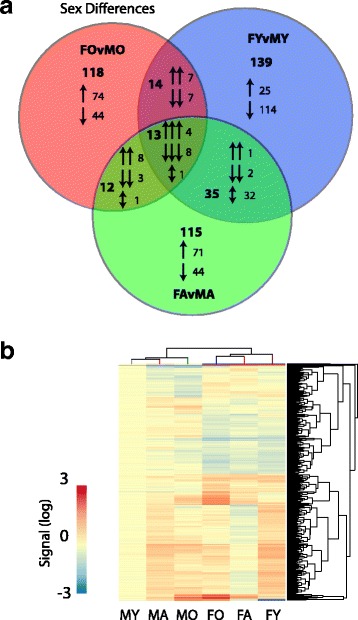
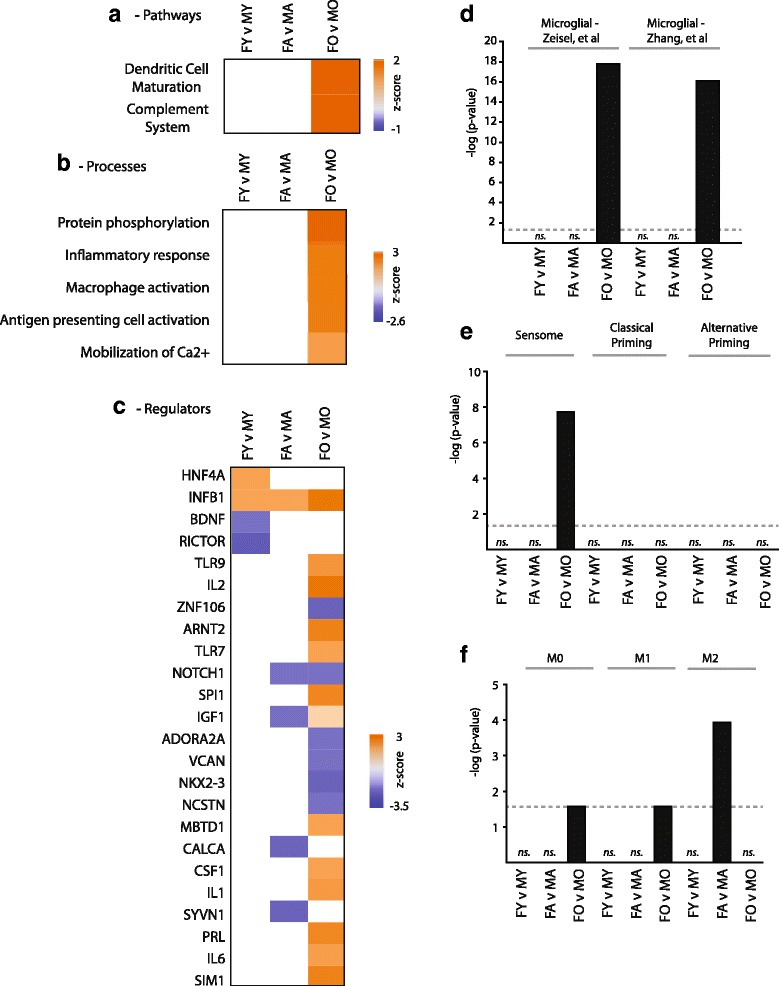
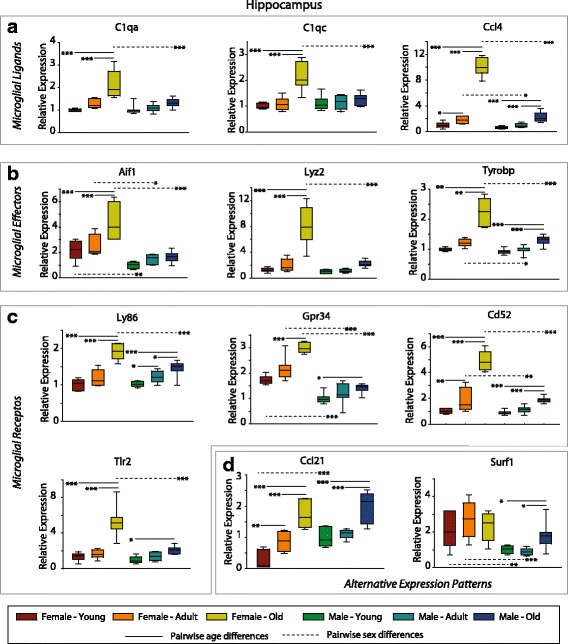
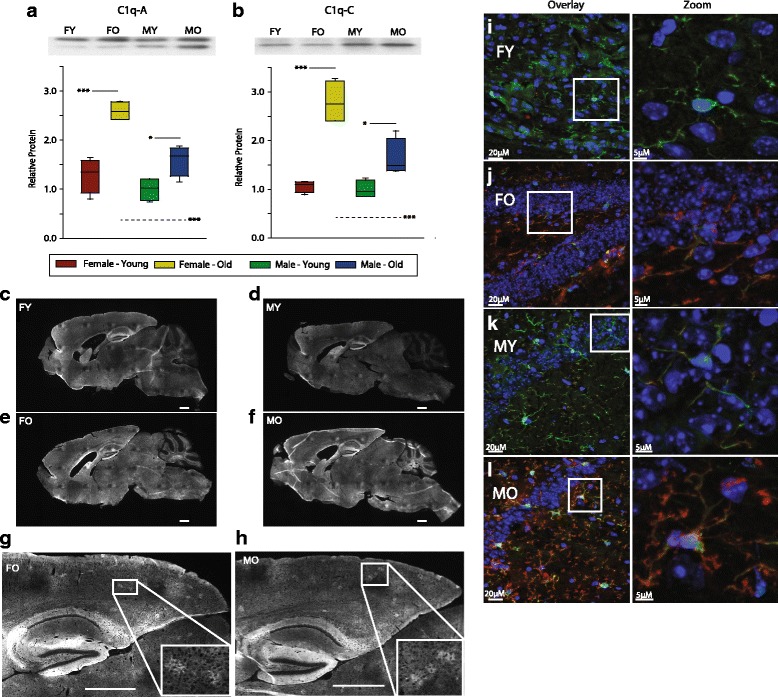
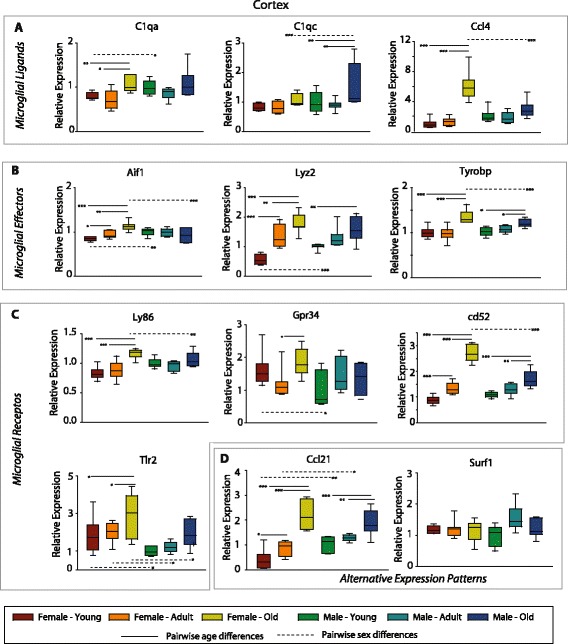
Results: Males and females demonstrate both common expression changes with aging and marked sex differences in the nature and magnitude of the aging responses. Age-related hippocampal induction of neuroinflammatory gene expression was sexually divergent and enriched for microglia-specific genes such as complement pathway components. Sexually divergent C1q protein expression was confirmed by immunoblotting and immunohistochemistry. Similar patterns of cortical sexually divergent gene expression were also evident. Additionally, inter-animal gene expression variability increased with aging in males, but not females.
Conclusions: These findings demonstrate sexually divergent neuroinflammation with aging that may contribute to sex differences in age-related neurological diseases such as stroke and Alzheimer's, specifically in the complement system. The increased expression variability in males suggests a loss of fidelity in gene expression regulation with aging. These findings reveal a central role of sex in the transcriptomic response of the hippocampus to aging that warrants further, in depth, investigations.
Keywords: Aging; Brain; Gene expression; Neuroinflammation; Sex differences.
Conflict of interest statement
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Berchtold NC, Cribbs DH, Coleman PD, Rogers J, Head E, Kim R, Beach T, Miller C, Troncoso J, Trojanowski JQ, et al. Gene expression changes in the course of normal brain aging are sexually dimorphic. Proc Natl Acad Sci U S A. 2008;105(40):15605–15610. doi: 10.1073/pnas.0806883105. - DOI - PMC - PubMed
-
- Blalock EM, Grondin R, Chen KC, Thibault O, Thibault V, Pandya JD, Dowling A, Zhang Z, Sullivan P, Porter NM, et al. Aging-related gene expression in hippocampus proper compared with dentate gyrus is selectively associated with metabolic syndrome variables in rhesus monkeys. J Neurosci. 2010;30(17):6058–6071. doi: 10.1523/JNEUROSCI.3956-09.2010. - DOI - PMC - PubMed
-
- Kadish I, Thibault O, Blalock EM, Chen KC, Gant JC, Porter NM, Landfield PW. Hippocampal and cognitive aging across the lifespan: a bioenergetic shift precedes and increased cholesterol trafficking parallels memory impairment. J Neurosci. 2009;29(6):1805–1816. doi: 10.1523/JNEUROSCI.4599-08.2009. - DOI - PMC - PubMed
-
- Masser DR, Bixler GV, Brucklacher RM, Yan H, Giles CB, Wren JD, Sonntag WE, Freeman WM. Hippocampal subregions exhibit both distinct and shared transcriptomic responses to aging and nonneurodegenerative cognitive decline. J Gerontol A Biol Sci Med Sci. 2014;69(11):1311–1324. doi: 10.1093/gerona/glu091. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
