

Improving Plant Nitrogen Use Efficiency through Alteration of Amino Acid Transport Processes
- PMID: 28733388
- PMCID: PMC5580756
- DOI: 10.1104/pp.17.00608
Improving Plant Nitrogen Use Efficiency through Alteration of Amino Acid Transport Processes
Abstract
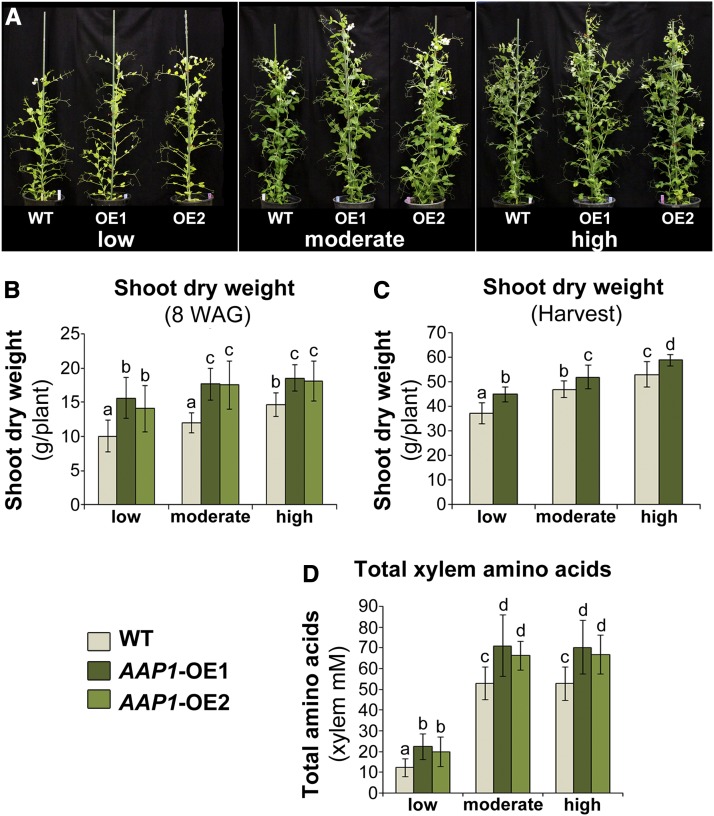
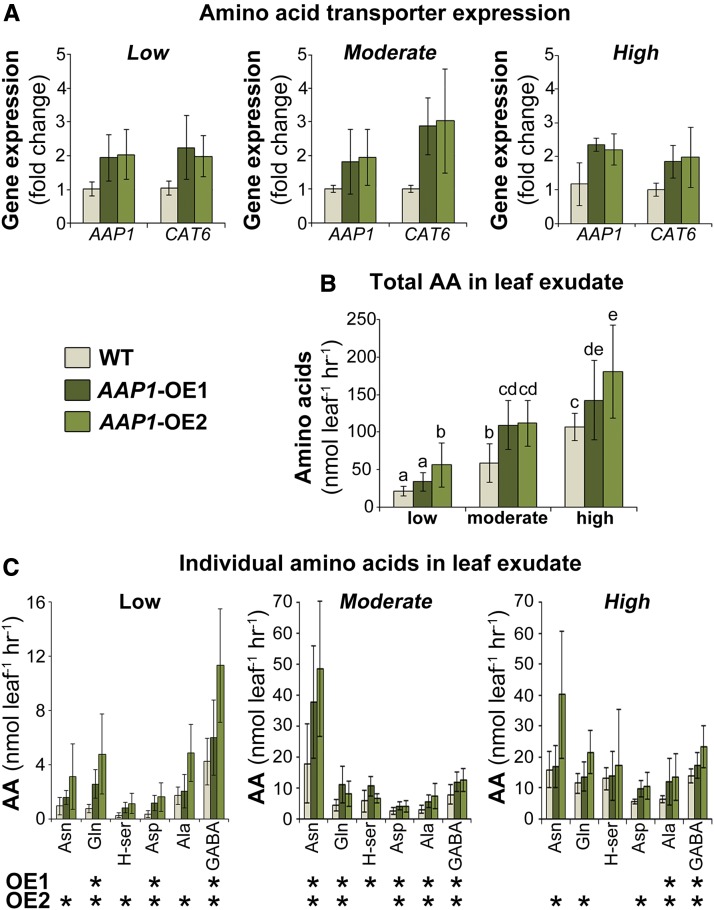
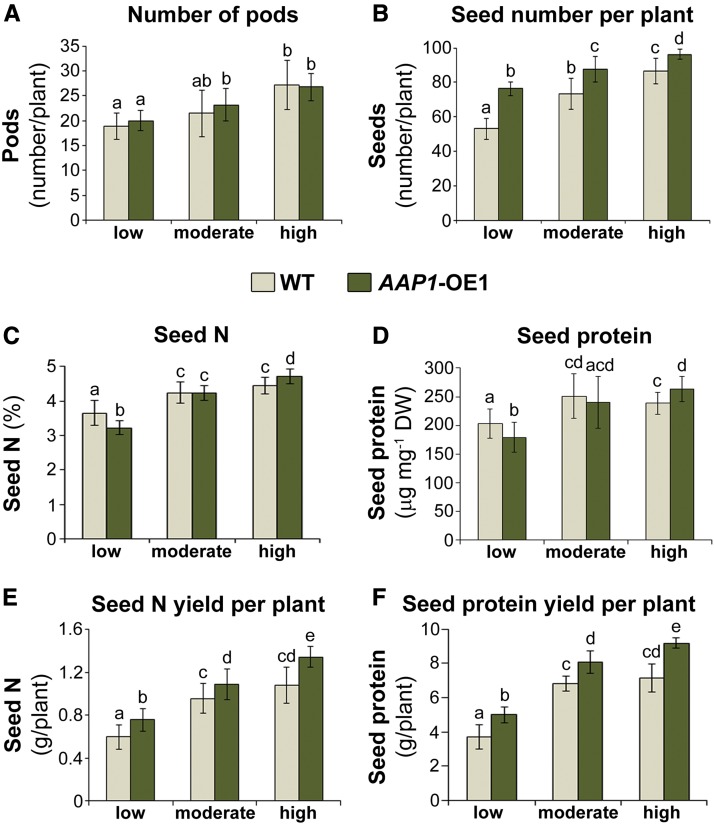
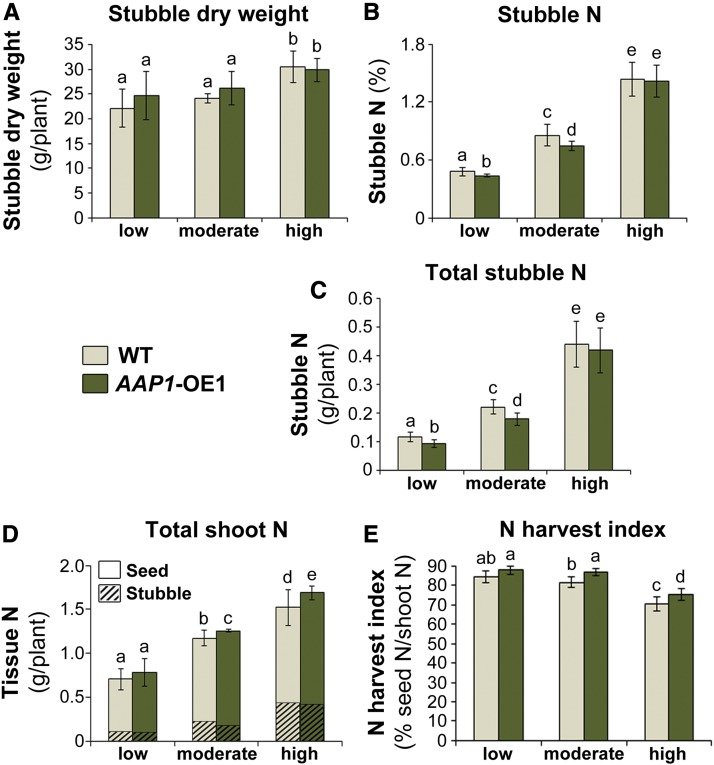
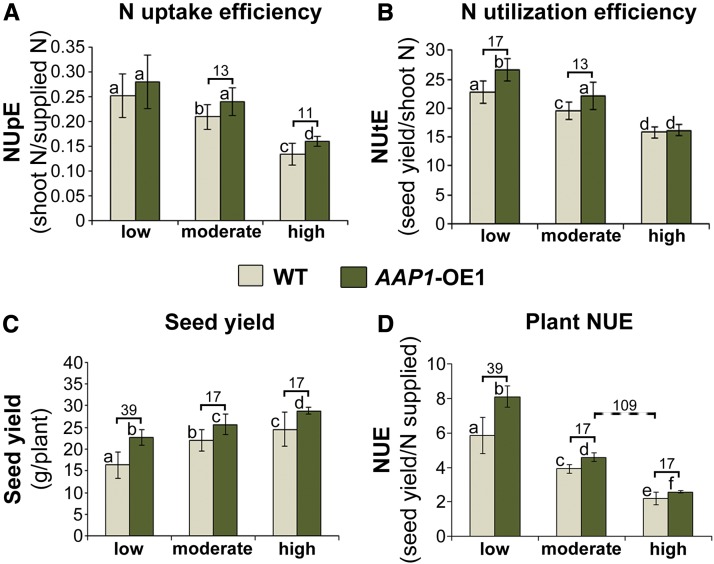
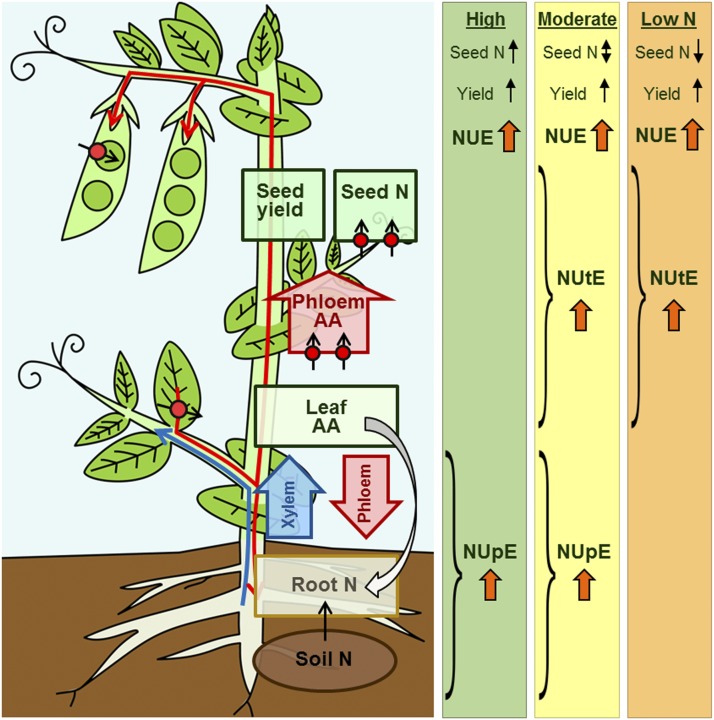
Improving the efficiency of nitrogen (N) uptake and utilization in plants could potentially increase crop yields while reducing N fertilization and, subsequently, environmental pollution. Within most plants, N is transported primarily as amino acids. In this study, pea (Pisum sativum) plants overexpressing AMINO ACID PERMEASE1 (AAP1) were used to determine if and how genetic manipulation of amino acid transport from source to sink affects plant N use efficiency. The modified plants were grown under low, moderate, or high N fertilization regimes. The results showed that, independent of the N nutrition, the engineered plants allocate more N via the vasculature to the shoot and seeds and produce more biomass and higher seed yields than wild-type plants. Dependent on the amount of N supplied, the AAP1-overexpressing plants displayed improved N uptake or utilization efficiency, or a combination of the two. They also showed significantly increased N use efficiency in N-deficient as well as in N-rich soils and, impressively, required half the amount of N to produce as many fruits and seeds as control plants. Together, these data support that engineering N allocation from source to sink presents an effective strategy to produce crop plants with improved productivity as well as N use efficiency in a range of N environments.
© 2017 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Ameziane R, Bernhard K, Lightfoot D (2000) Expression of the bacterial gdhA gene encoding a NADPH glutamate dehydrogenase in tobacco affects plant growth and development. Plant Soil 22: 147–157
-
- Aoyama C, Santa T, Tsunoda M, Fukushima T, Kitada C, Imai K (2004) A fully automated amino acid analyzer using NBD-F as a fluorescent derivatization reagent. Biomed Chromatogr 18: 630–636 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
