

Genome Organization Drives Chromosome Fragility
- PMID: 28735753
- PMCID: PMC6133249
- DOI: 10.1016/j.cell.2017.06.034
Genome Organization Drives Chromosome Fragility
Abstract
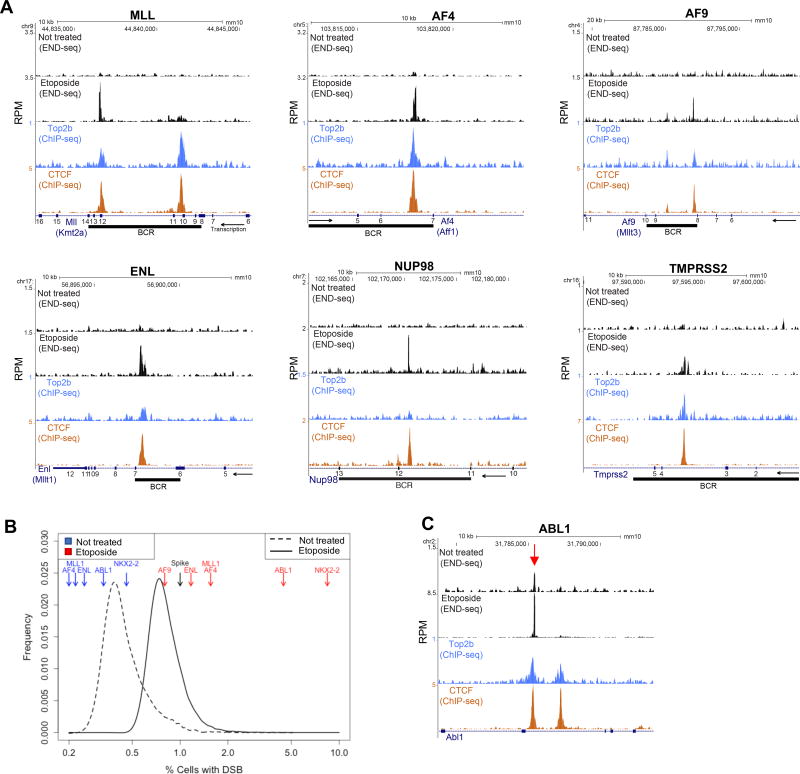
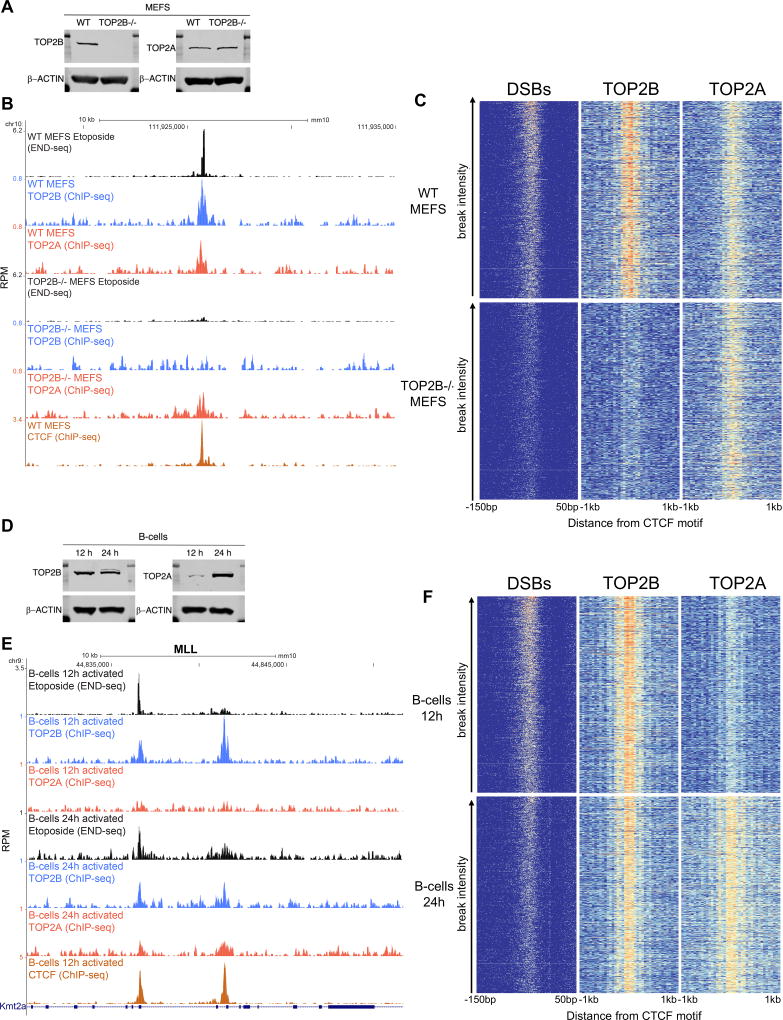
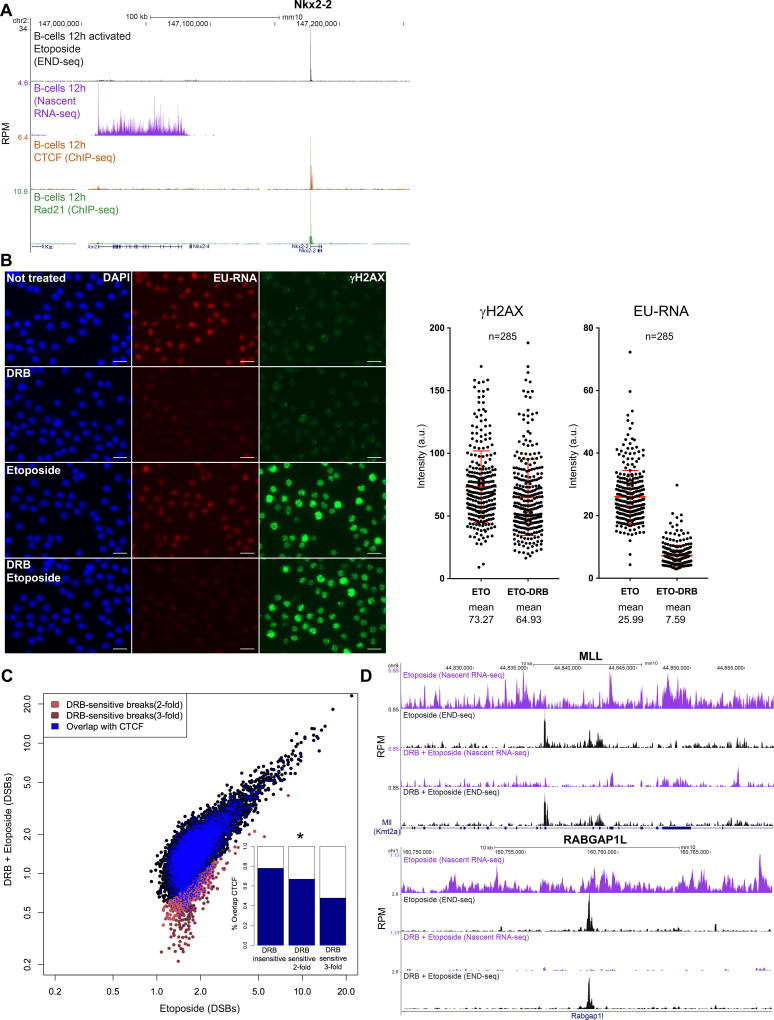
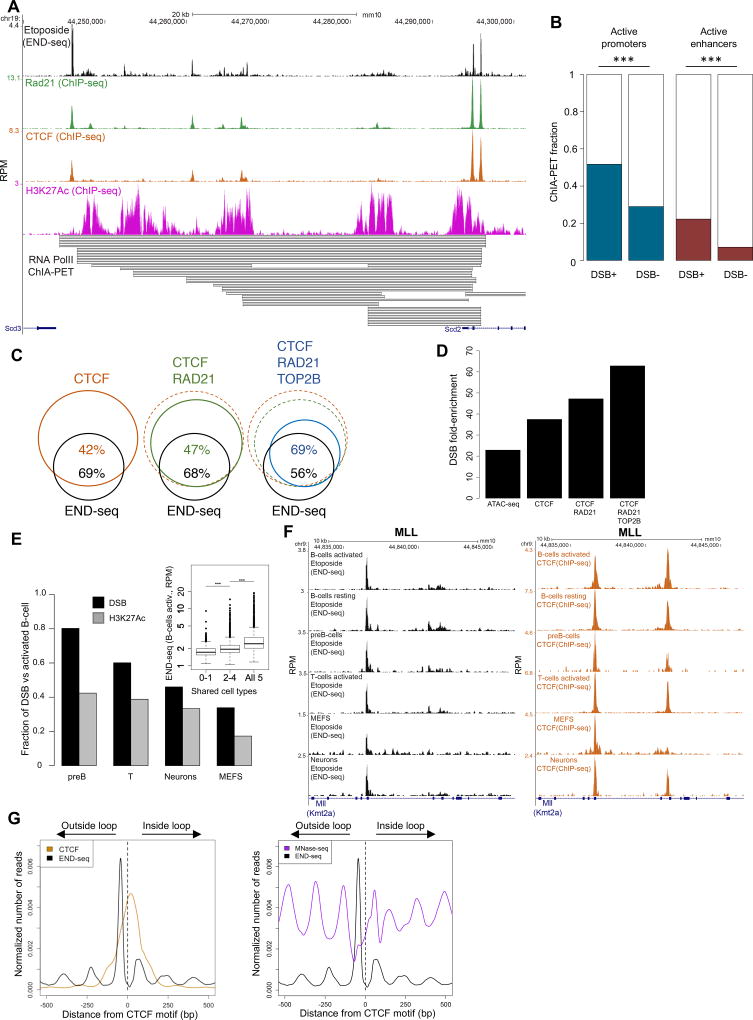
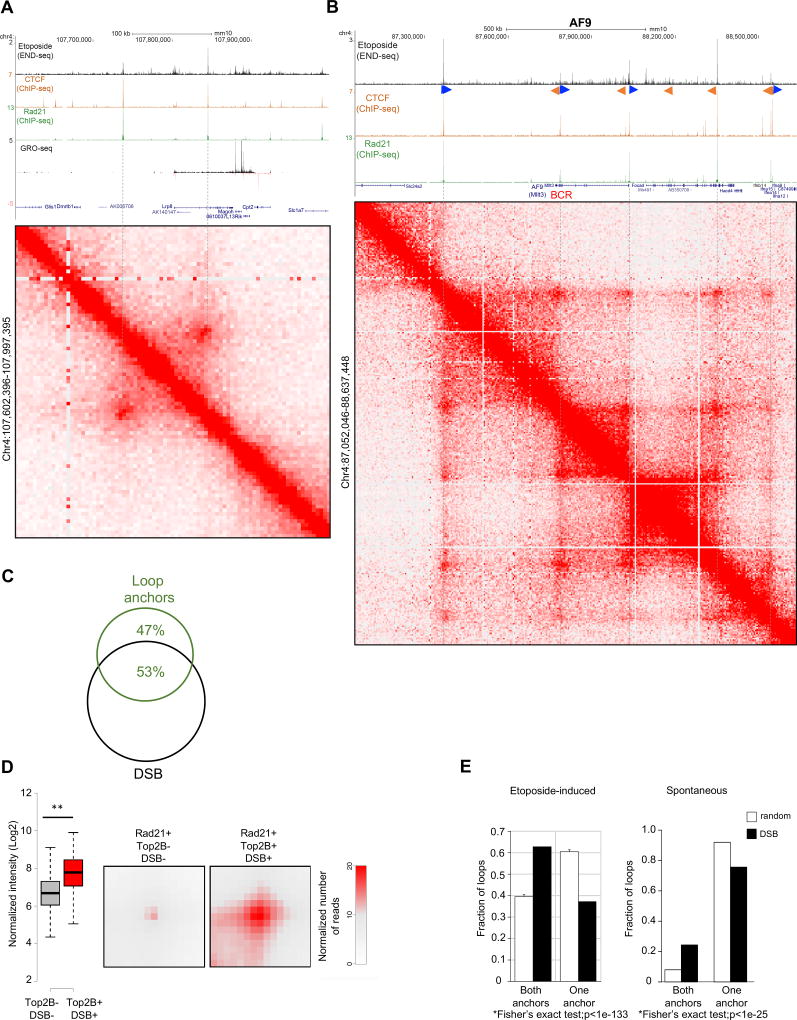
In this study, we show that evolutionarily conserved chromosome loop anchors bound by CCCTC-binding factor (CTCF) and cohesin are vulnerable to DNA double strand breaks (DSBs) mediated by topoisomerase 2B (TOP2B). Polymorphisms in the genome that redistribute CTCF/cohesin occupancy rewire DNA cleavage sites to novel loop anchors. While transcription- and replication-coupled genomic rearrangements have been well documented, we demonstrate that DSBs formed at loop anchors are largely transcription-, replication-, and cell-type-independent. DSBs are continuously formed throughout interphase, are enriched on both sides of strong topological domain borders, and frequently occur at breakpoint clusters commonly translocated in cancer. Thus, loop anchors serve as fragile sites that generate DSBs and chromosomal rearrangements. VIDEO ABSTRACT.
Keywords: DNA breaks; breakpoint cluster regions; cancer; fragile sites; genome instability; topoisomerase; topologically associated domains; translocations.
Published by Elsevier Inc.
Figures

Comment in
-
Anchoring Chromatin Loops to Cancer.Dev Cell. 2017 Aug 7;42(3):209-211. doi: 10.1016/j.devcel.2017.07.013. Dev Cell. 2017. PMID: 28787588 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
