

β-TrCP1 Is a Vacillatory Regulator of Wnt Signaling
- PMID: 28736239
- PMCID: PMC5634333
- DOI: 10.1016/j.chembiol.2017.06.009
β-TrCP1 Is a Vacillatory Regulator of Wnt Signaling
Abstract
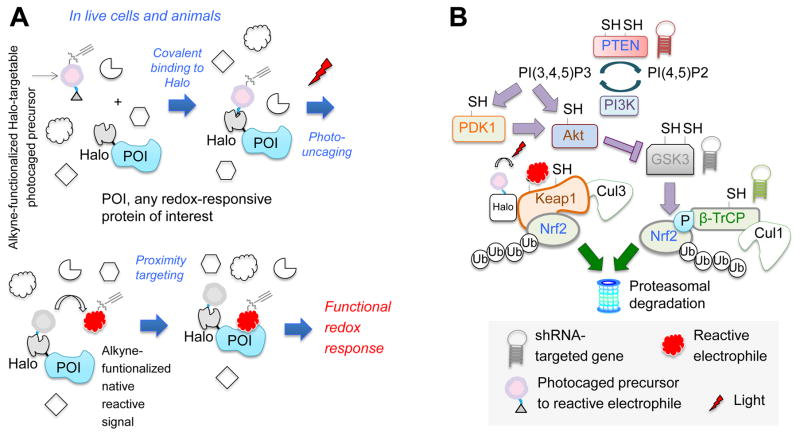
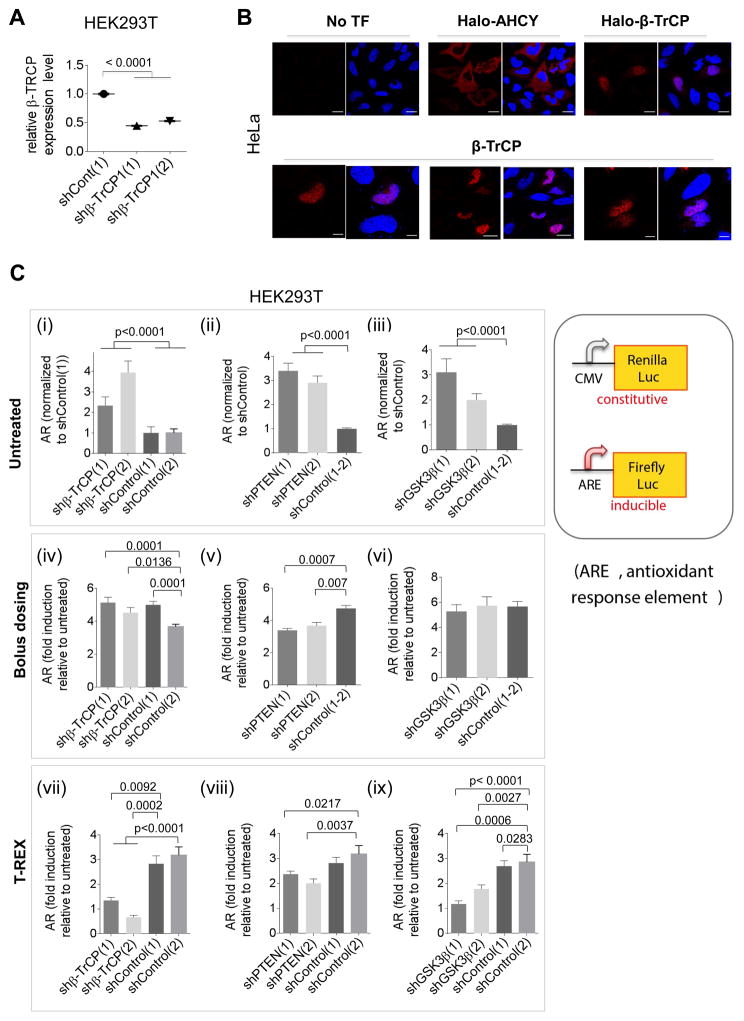
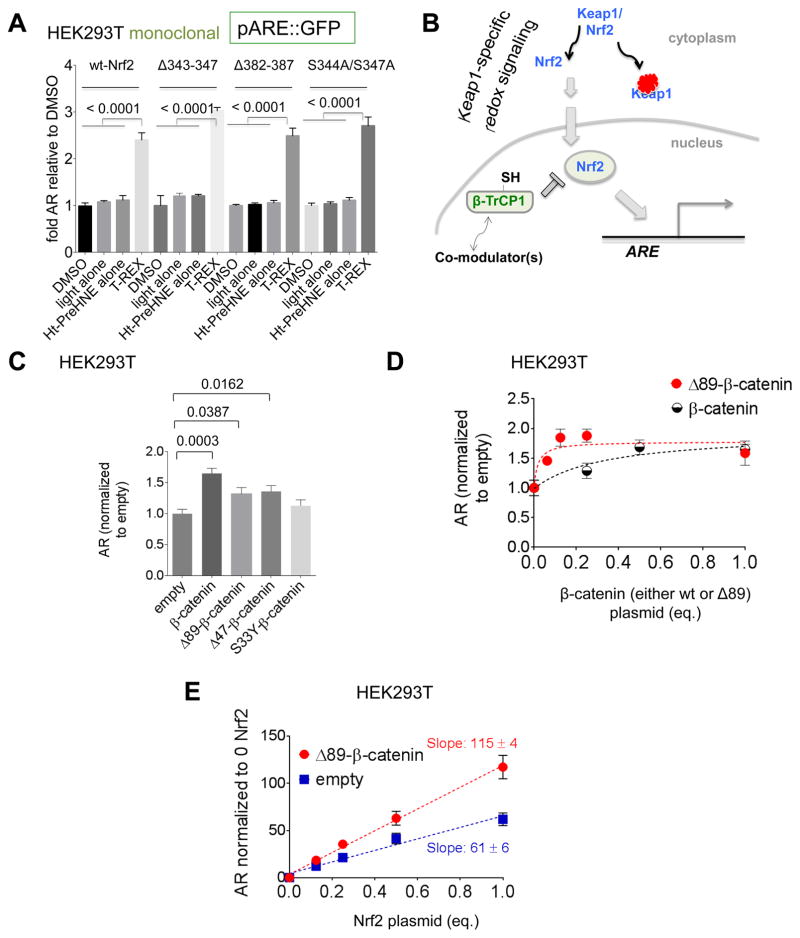
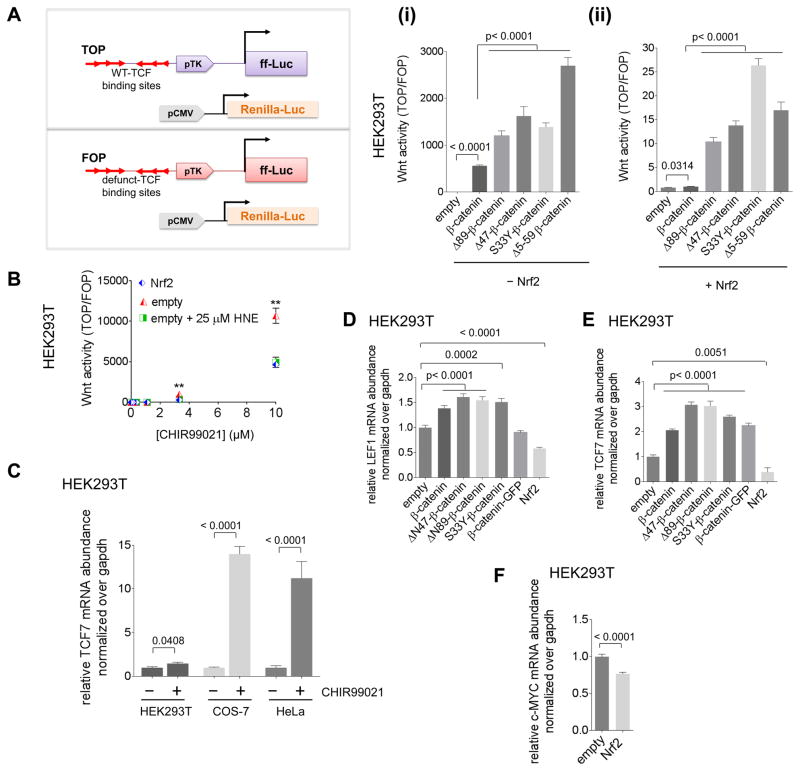
Simultaneous hyperactivation of Wnt and antioxidant response (AR) are often observed during oncogenesis. However, it remains unclear how the β-catenin-driven Wnt and the Nrf2-driven AR mutually regulate each other. The situation is compounded because many players in these two pathways are redox sensors, rendering bolus redox signal-dosing methods uninformative. Herein we examine the ramifications of single-protein target-specific AR upregulation in various knockdown lines. Our data document that Nrf2/AR strongly inhibits β-catenin/Wnt. The magnitude and mechanism of this negative regulation are dependent on the direct interaction between β-catenin N terminus and β-TrCP1 (an antagonist of both Nrf2 and β-catenin), and independent of binding between Nrf2 and β-TrCP1. Intriguingly, β-catenin positively regulates AR. Because AR is a negative regulator of Wnt regardless of β-catenin N terminus, this switch of function is likely sufficient to establish a new Wnt/AR equilibrium during tumorigenesis.
Keywords: 4-hydroxynonenal; HaloTag; Keap1-Nrf2-antioxidant response; reactive electrophile response; redox signaling; signaling crosstalk; β-TrCP; β-catenin/wnt signaling.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
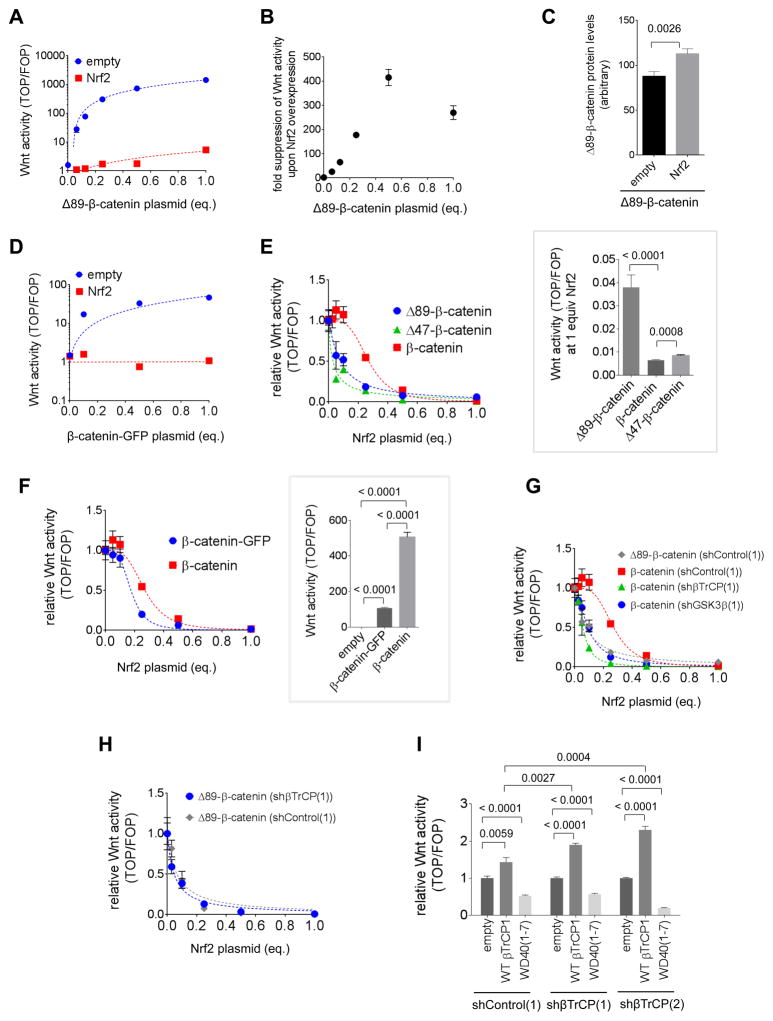
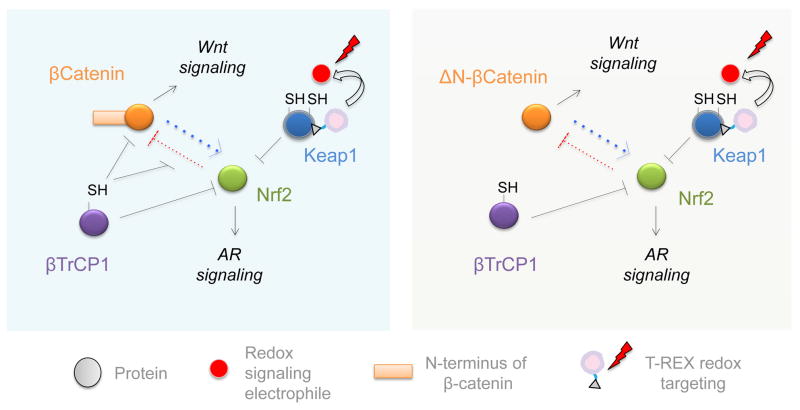
References
-
- Alomar SY, Mansour L, Abuderman A, Alkhuriji A, Arafah M, Alwasel S, Harrath AH, Almutairi M, Trayhyrn P, Dar JA. β-Catenin accumulation and S33F mutation of CTNNB1 gene in colorectal cancer in Saudi Arabia. Polish Journal of Pathology. 2016;2:156–162. - PubMed
-
- Bryan HK, Olayanju A, Goldring CE, Park BK. The Nrf2 cell defence pathway: Keap1-dependent and -independent mechanisms of regulation. Biochemical pharmacology. 2013;85:705–717. - PubMed
-
- Cadigan KM, Nusse R. Wnt signaling: a common theme in animal development. Genes & development. 1997;11:3286–3305. - PubMed
-
- Chan E, Gat U, McNiff JM, Fuchs E. A common human skin tumour is caused by activating mutations in [beta]-catenin. Nat Genet. 1999;21:410–413. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
