

Anion-selective Formate/nitrite transporters: taxonomic distribution, phylogenetic analysis and subfamily-specific conservation pattern in prokaryotes
- PMID: 28738779
- PMCID: PMC5525234
- DOI: 10.1186/s12864-017-3947-4
Anion-selective Formate/nitrite transporters: taxonomic distribution, phylogenetic analysis and subfamily-specific conservation pattern in prokaryotes
Abstract
Background: The monovalent anions formate, nitrite and hydrosulphide are main metabolites of bacterial respiration during anaerobic mixed-acid fermentation. When accumulated in the cytoplasm, these anions become cytotoxic. Membrane proteins that selectively transport these monovalent anions across the membrane have been identified and they belong to the family of Formate/Nitrite Transporters (FNTs). Individual members that selectively transport formate, nitrite and hydrosulphide have been investigated. Experimentally determined structures of FNTs indicate that they share the same hourglass helical fold with aquaporins and aquaglyceroporins and have two constriction regions, namely, cytoplasmic slit and central constriction. Members of FNTs are found in bacteria, archaea, fungi and protists. However, no FNT homolog has been identified in mammals. With FNTs as potential drug targets for many bacterial diseases, it is important to understand the mechanism of selectivity and transport across these transporters.
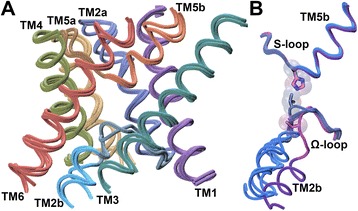
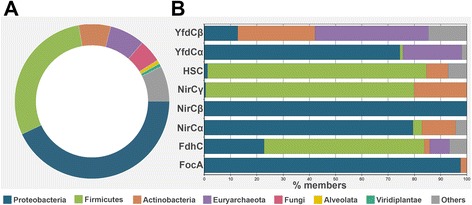
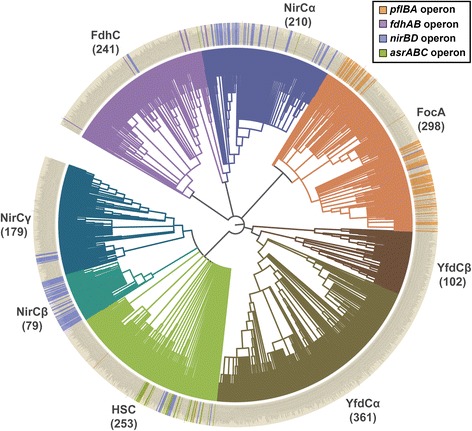
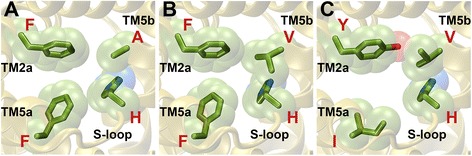
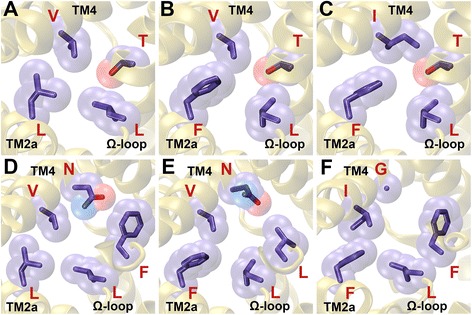
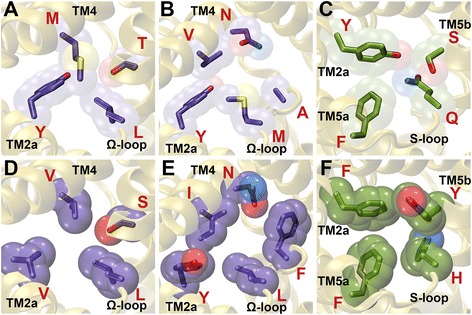
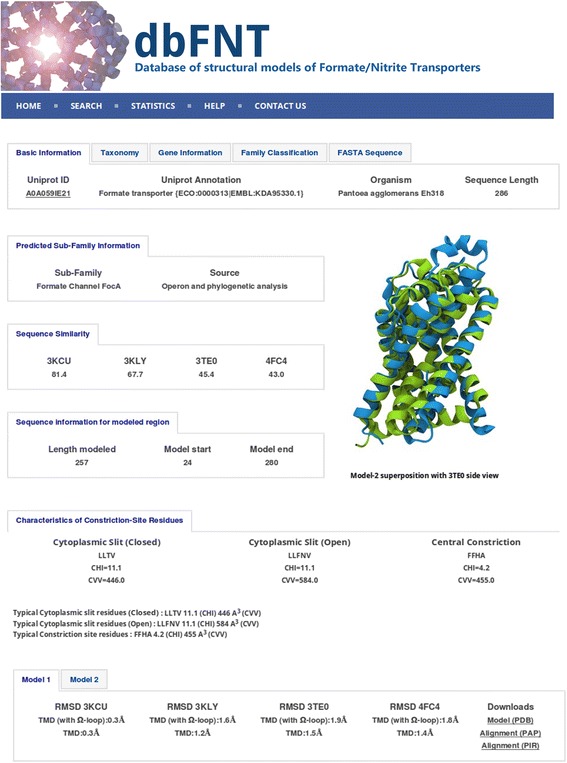
Results: We have systematically searched the sequence databases and identified 2206 FNT sequences from bacteria, archaea and eukaryotes. Although FNT sequences are very diverse, homology modeling followed by structure-based sequence alignment revealed that nearly one third of all the positions within the transmembrane region exhibit high conservation either as a group or at the level of individual residues across all three kingdoms. Phylogenetic analysis of prokaryotic FNT sequences revealed eight different subgroups. Formate, nitrite and hydrosulphide transporters respectively are clustered into two (FocA and FdhC), three (NirC-α, NirC-β and NirC-γ) and one (HSC) subfamilies. We have also recognized two FNT subgroups (YfdC-α and YfdC-β) with unassigned function. Analysis of taxonomic distribution indicates that each subfamily prefers specific taxonomic groups. Structure-based sequence alignment of individual subfamily members revealed that certain positions in the two constriction regions and some residues facing the interior show subfamily-specific conservation. We have also identified examples of FNTs with the two constriction regions formed by residues that are less frequently observed. We have developed dbFNT, a database of FNT models and associated details. dbFNT is freely available to scientific community.
Conclusions: Taxonomic distribution and sequence conservation of FNTs exhibit subfamily-specific features. The conservation pattern in the central constriction and cytoplasmic slit in the open and closed states are distinct for YfdC and NirC subfamilies. The same is true for some residues facing the interior of the transporters. The specific residues in these positions can exert influence on the type of solutes that are transported by these proteins. With FNTs found in many disease-causing bacteria, the knowledge gained in this study can be used in the development and design of anti-bacterial drugs.
Keywords: Anion transport; Channel selectivity and transport; Conformational changes; FNT family; Gating; Homology modeling; Hour-glass helical fold; Phylogenetic analysis; Sequence diversity; Structure-based sequence alignment.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
Not applicable.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Oren A. Anaerobic respiration. Chichester: John Wiley & Sons Ltd.; 2009.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
