

Two functionally distinct E2/E3 pairs coordinate sequential ubiquitination of a common substrate in Caenorhabditis elegans development
- PMID: 28739890
- PMCID: PMC5559030
- DOI: 10.1073/pnas.1705060114
Two functionally distinct E2/E3 pairs coordinate sequential ubiquitination of a common substrate in Caenorhabditis elegans development
Abstract
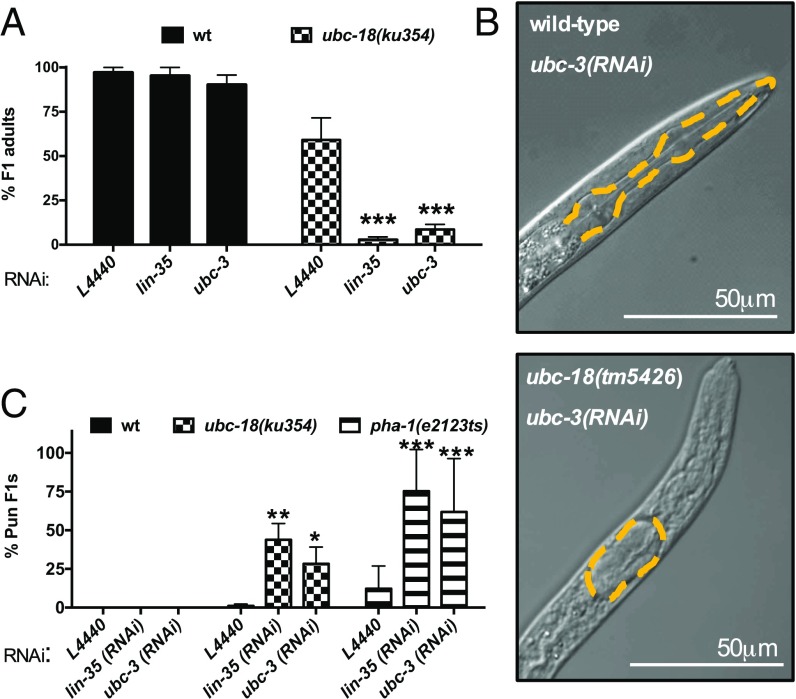
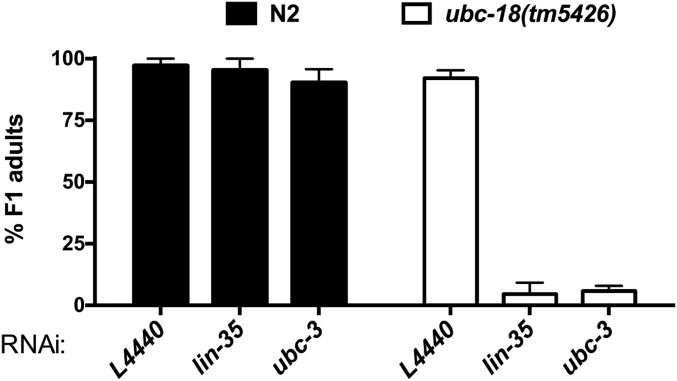
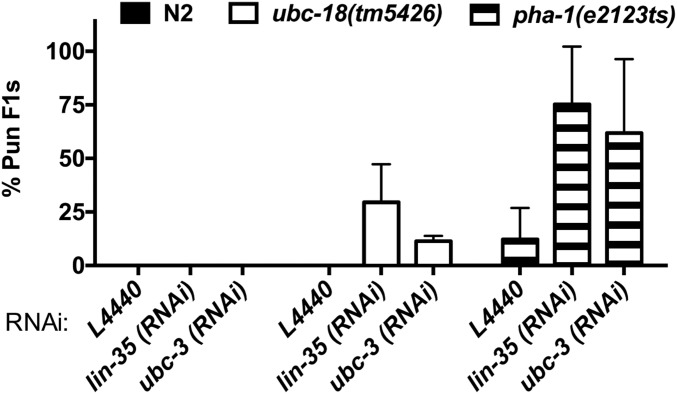
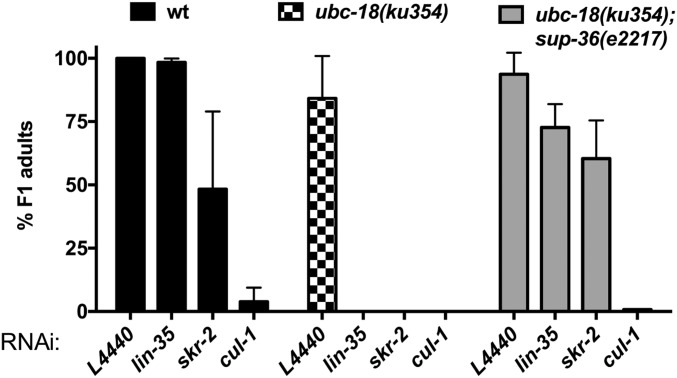
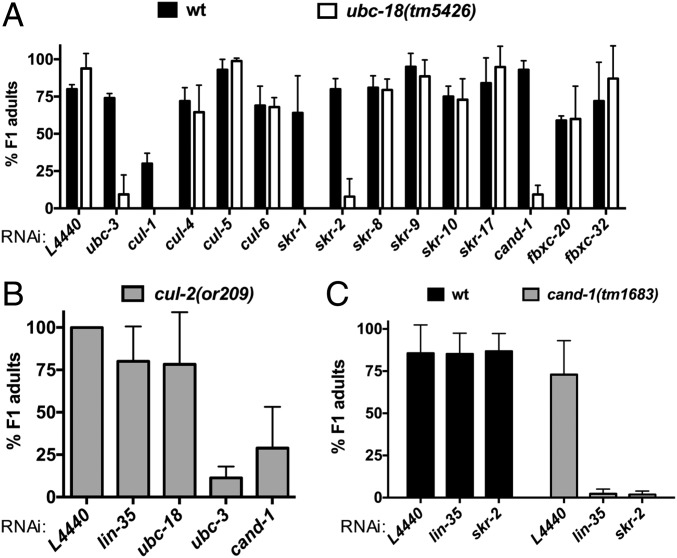
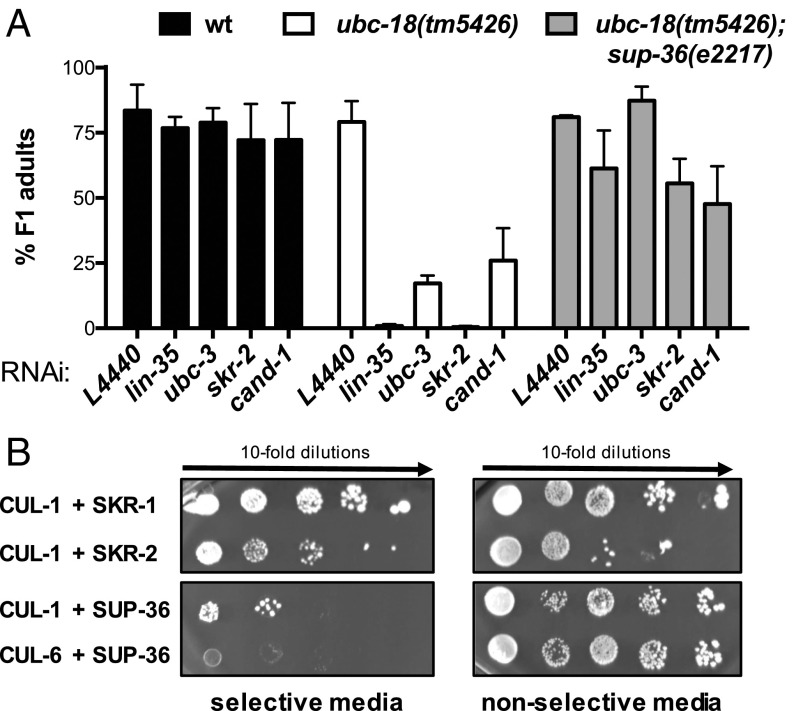
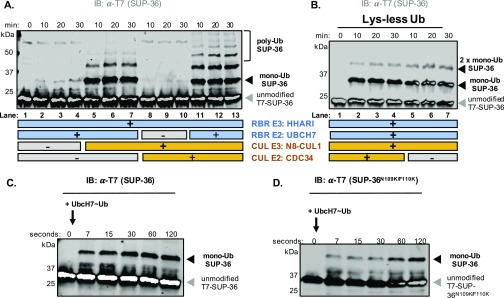
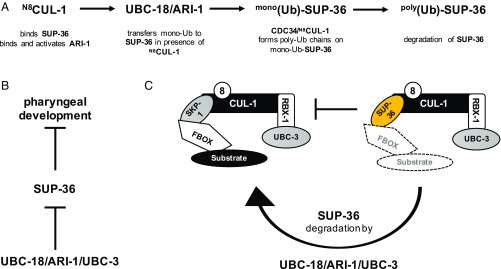
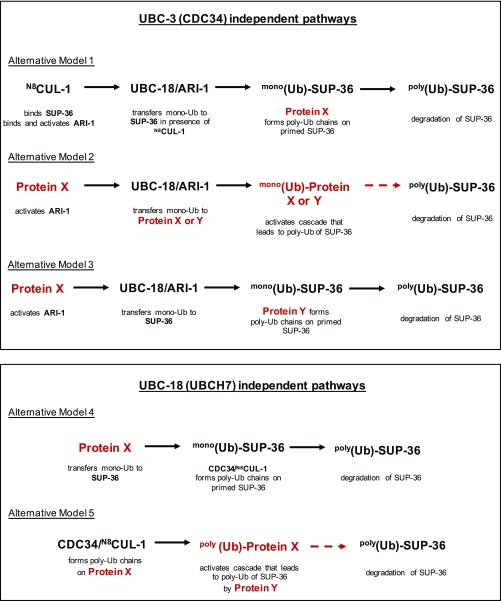
Ubiquitination, the crucial posttranslational modification that regulates the eukaryotic proteome, is carried out by a trio of enzymes, known as E1 [ubiquitin (Ub)-activating enzyme], E2 (Ub-conjugating enzyme), and E3 (Ub ligase). Although most E2s can work with any of the three mechanistically distinct classes of E3s, the E2 UBCH7 is unable to function with really interesting new gene (RING)-type E3s, thereby restricting it to homologous to E6AP C-terminus (HECT) and RING-in-between-RING (RBR) E3s. The Caenorhabditis elegans UBCH7 homolog, UBC-18, plays a critical role in developmental processes through its cooperation with the RBR E3 ARI-1 (HHARI in humans). We discovered that another E2, ubc-3, interacts genetically with ubc-18 in an unbiased genome-wide RNAi screen in C. elegans These two E2s have nonoverlapping biochemical activities, and each is dedicated to distinct classes of E3s. UBC-3 is the ortholog of CDC34 that functions specifically with Cullin-RING E3 ligases, such as SCF (Skp1-Cullin-F-box). Our genetic and biochemical studies show that UBCH7 (UBC-18) and the RBR E3 HHARI (ARI-1) coordinate with CDC34 (UBC-3) and an SCF E3 complex to ubiquitinate a common substrate, a SKP1-related protein. We show that UBCH7/HHARI primes the substrate with a single Ub in the presence of CUL-1, and that CDC34 is required to build chains onto the Ub-primed substrate. Our study reveals that the association and coordination of two distinct E2/E3 pairs play essential roles in a developmental pathway and suggests that cooperative action among E3s is a conserved feature from worms to humans.
Keywords: CDC34; Cullin-RING ligase; RBR E3 ligase; UBCH7; ubiquitin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Schnell JD, Hicke L. Non-traditional functions of ubiquitin and ubiquitin-binding proteins. J Biol Chem. 2003;278:35857–35860. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
