

The Role of Inflammation in β-cell Dedifferentiation
- PMID: 28740254
- PMCID: PMC5524956
- DOI: 10.1038/s41598-017-06731-w
The Role of Inflammation in β-cell Dedifferentiation
Abstract
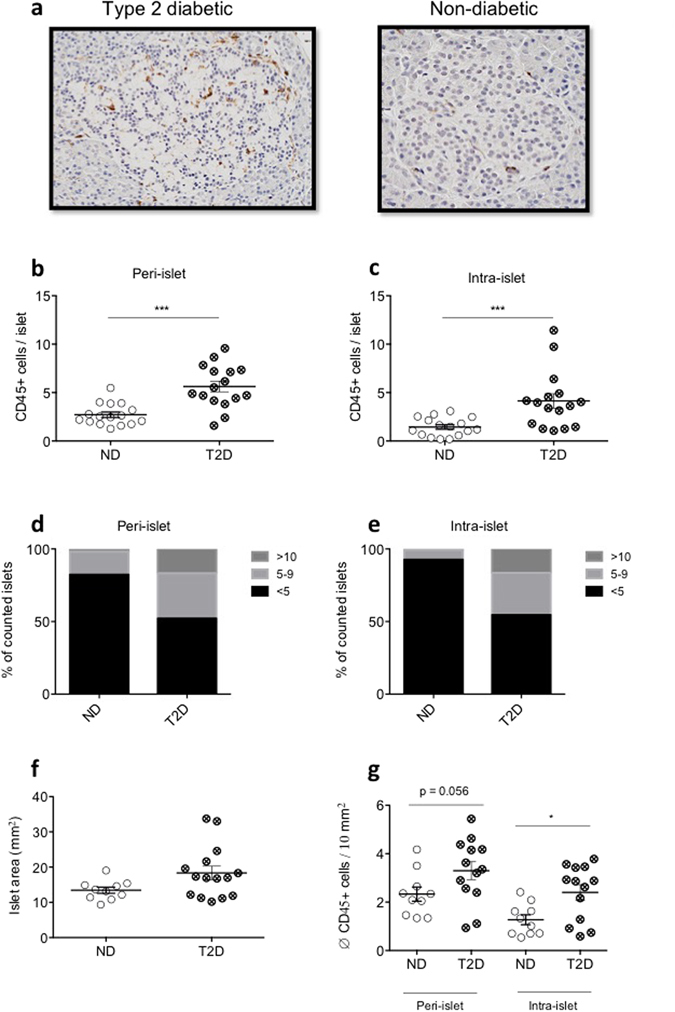
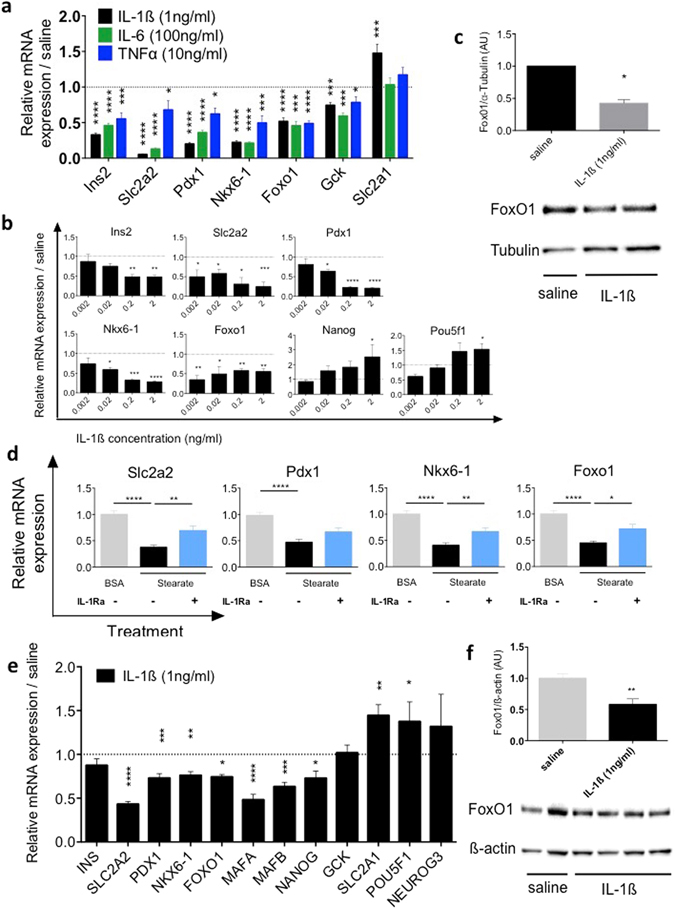
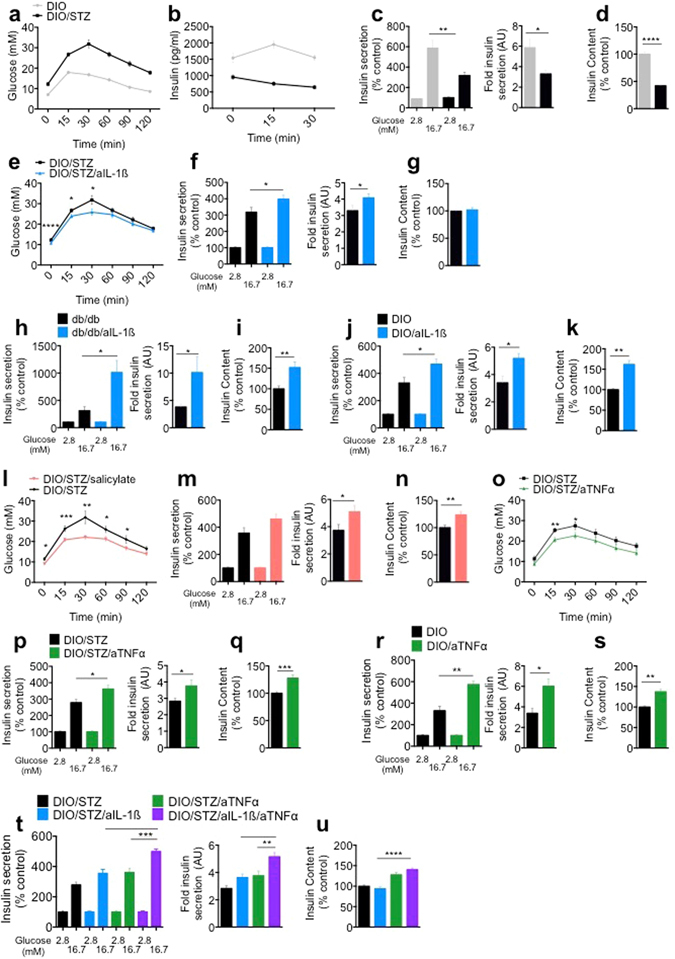
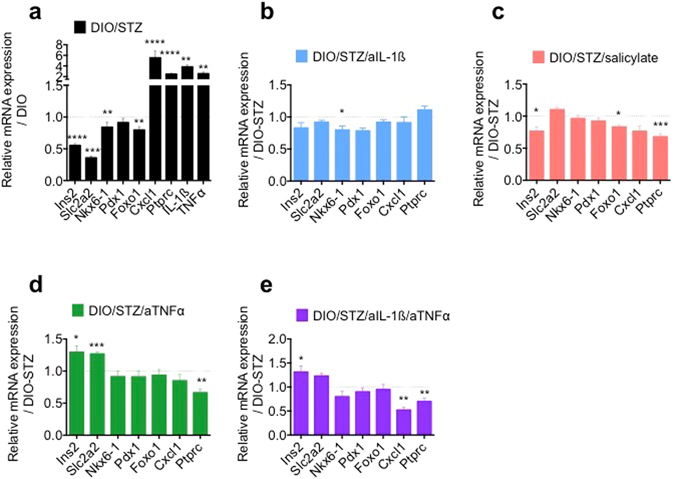
Chronic inflammation impairs insulin secretion and sensitivity. β-cell dedifferentiation has recently been proposed as a mechanism underlying β-cell failure in T2D. Yet the effect of inflammation on β-cell identity in T2D has not been studied. Therefore, we investigated whether pro-inflammatory cytokines induce β-cell dedifferentiation and whether anti-inflammatory treatments improve insulin secretion via β-cell redifferentiation. We observed that IL-1β, IL-6 and TNFα promote β-cell dedifferentiation in cultured human and mouse islets, with IL-1β being the most potent one of them. In particular, β-cell identity maintaining transcription factor Foxo1 was downregulated upon IL-1β exposure. In vivo, anti-IL-1β, anti-TNFα or NF-kB inhibiting sodium salicylate treatment improved insulin secretion of isolated islets. However, only TNFα antagonism partially prevented the loss of β-cell identity gene expression. Finally, the combination of IL-1β and TNFα antagonism improved insulin secretion of ex vivo isolated islets in a synergistic manner. Thus, while inflammation triggered β-cell dedifferentiation and dysfunction in vitro, this mechanism seems to be only partly responsible for the observed in vivo improvements in insulin secretion.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
