

NADPH oxidase 4 is required for the generation of macrophage migration inhibitory factor and host defense against Toxoplasma gondii infection
- PMID: 28743960
- PMCID: PMC5526938
- DOI: 10.1038/s41598-017-06610-4
NADPH oxidase 4 is required for the generation of macrophage migration inhibitory factor and host defense against Toxoplasma gondii infection
Abstract
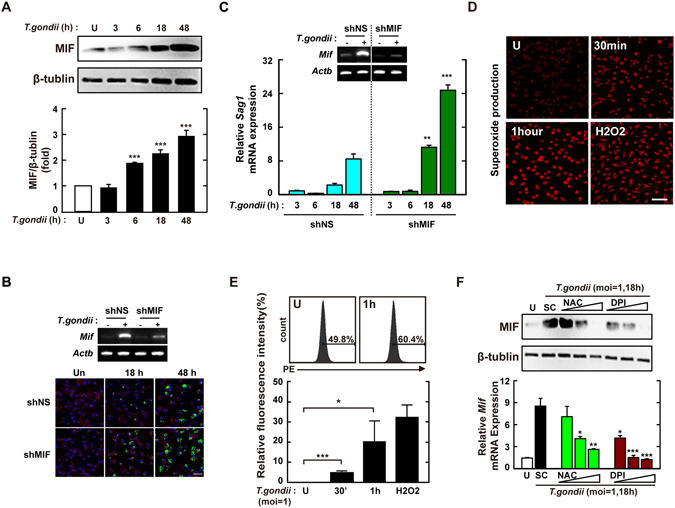
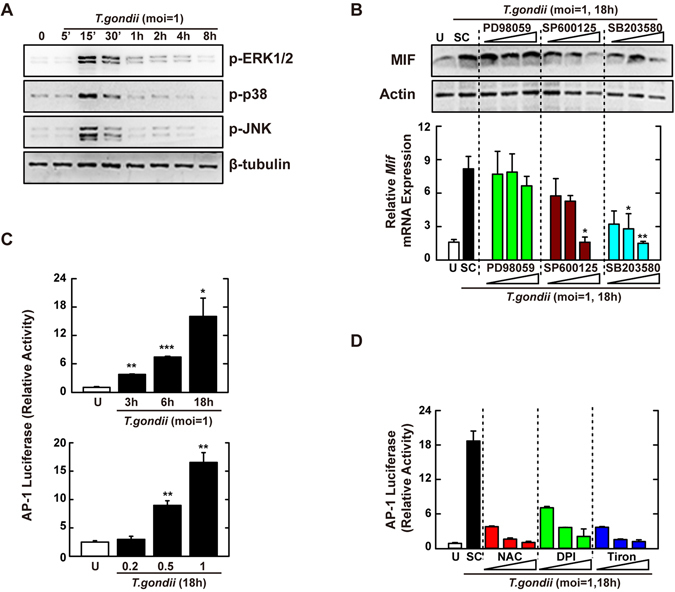
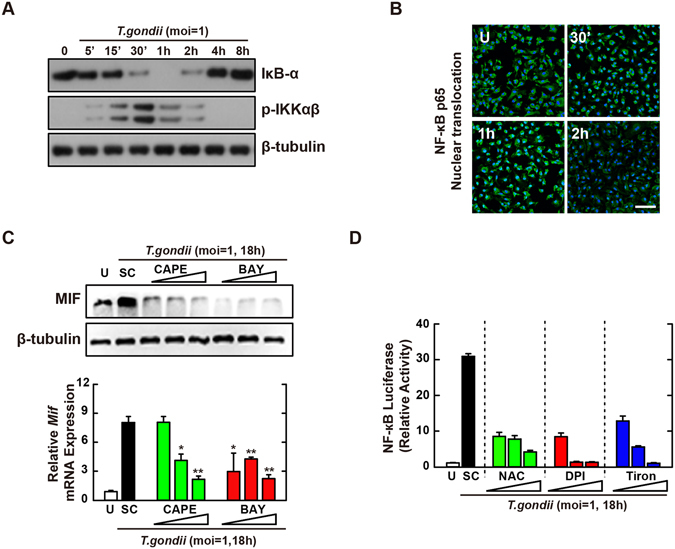
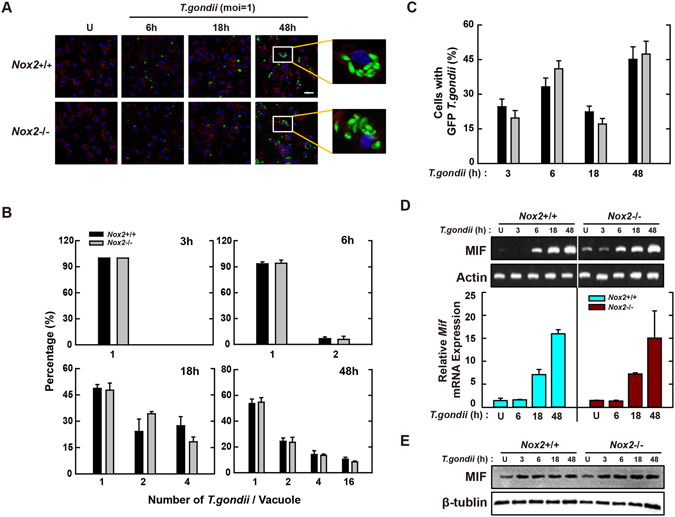
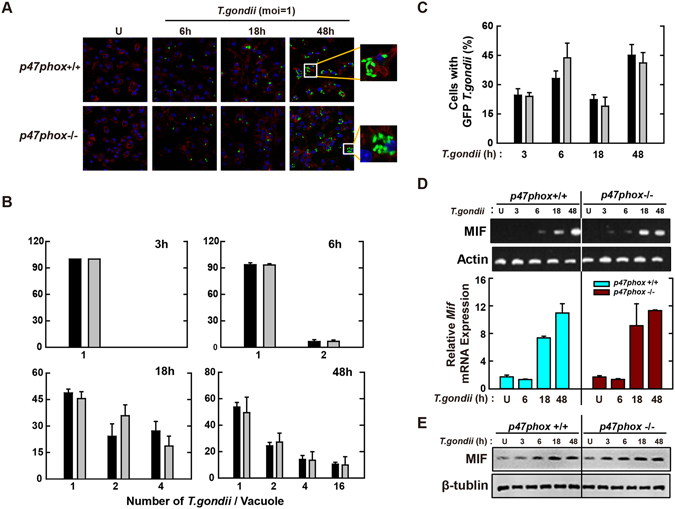
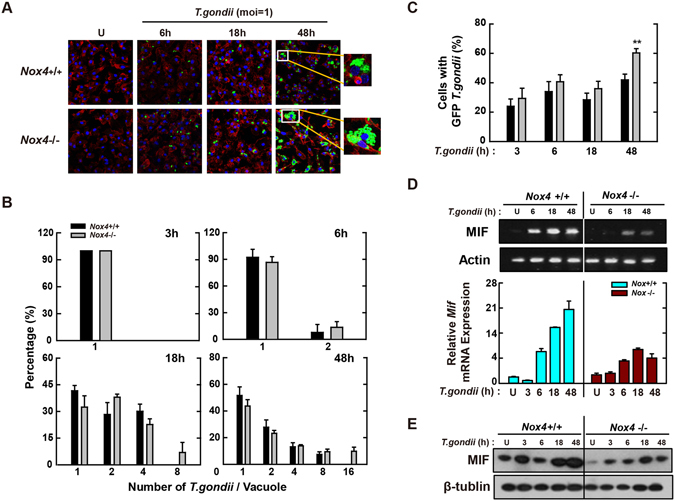
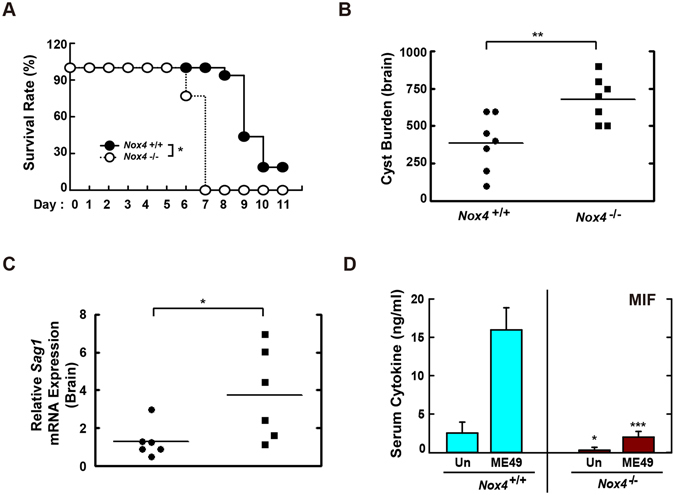
Nicotinamide adenine dinucleotide phosphate (NADPH) oxidases (Nox) are an important family of catalytic enzymes that generate reactive oxygen species (ROS), which mediate the regulation of diverse cellular functions. Although phagocyte Nox2/gp91phox is closely associated with the activation of host innate immune responses, the roles of Nox family protein during Toxoplasma gondii (T. gondii) infection have not been fully investigated. Here, we found that T. gondii-mediated ROS production was required for the upregulation of macrophage migration inhibitory factor (MIF) mRNA and protein levels via activation of mitogen-activated protein kinase and nuclear factor-κB signaling in macrophages. Interestingly, MIF knockdown led to a significant increase in the survival of intracellular T. gondii in bone marrow-derived macrophages (BMDMs). Moreover, Nox4 deficiency, but not Nox2/gp91phox and the cytosolic subunit p47phox, resulted in enhanced survival of the intracellular T. gondii RH strain and impaired expression of T. gondii-mediated MIF in BMDMs. Additionally, Nox4-deficient mice showed increased susceptibility to virulent RH strain infection and increased cyst burden in brain tissues and low levels of MIF expression following infection with the avirulent ME49 strain. Collectively, our findings indicate that Nox4-mediated ROS generation plays a central role in MIF production and resistance to T. gondii infection.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
