

New Challenges to Study Heterogeneity in Cancer Redox Metabolism
- PMID: 28744456
- PMCID: PMC5504267
- DOI: 10.3389/fcell.2017.00065
New Challenges to Study Heterogeneity in Cancer Redox Metabolism
Abstract
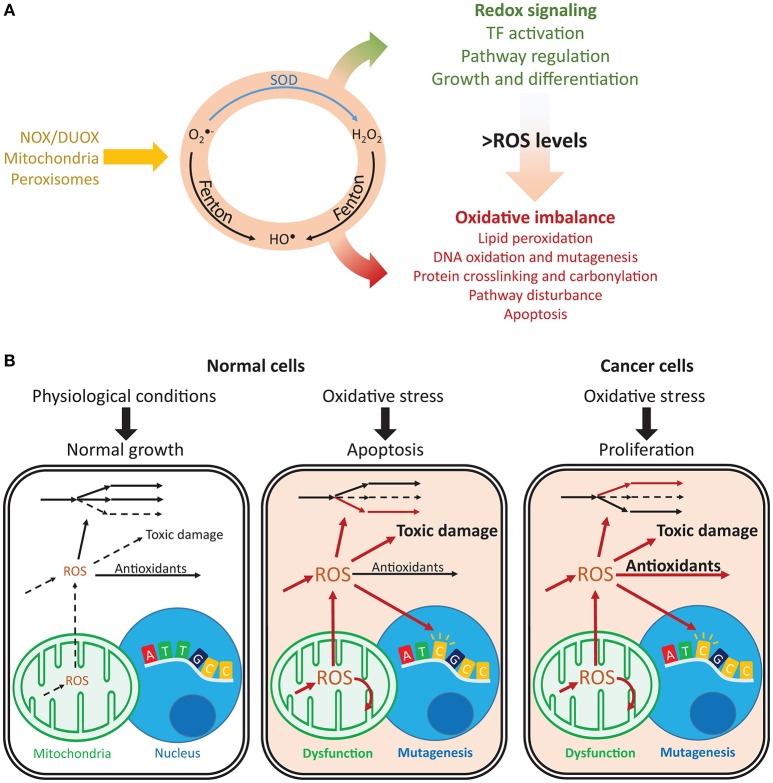
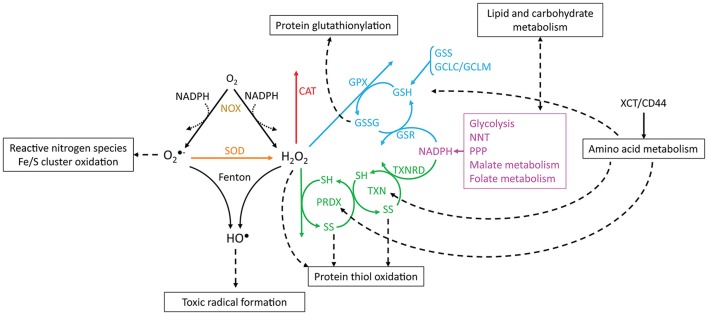
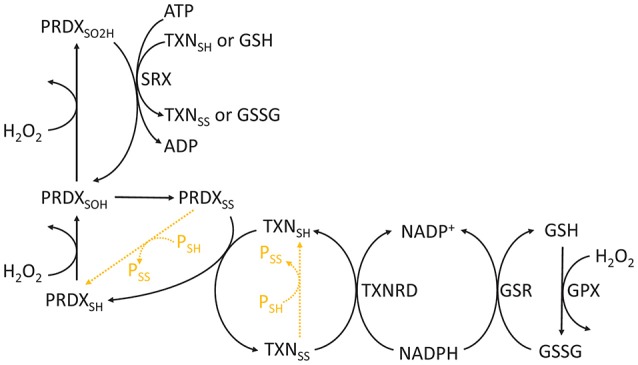
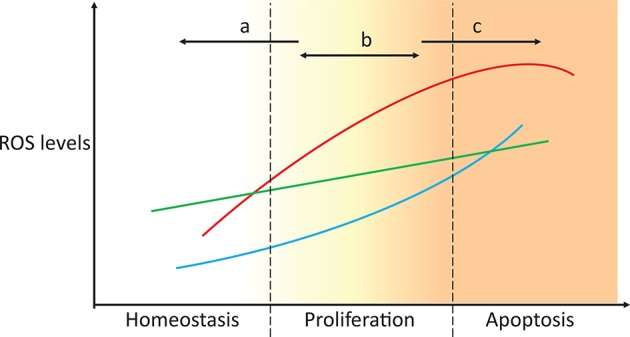
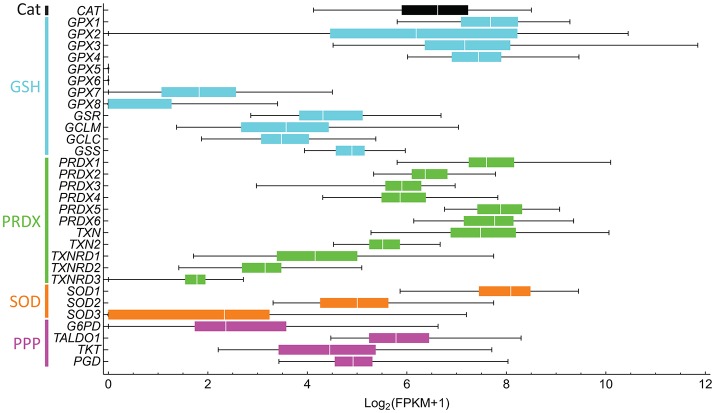
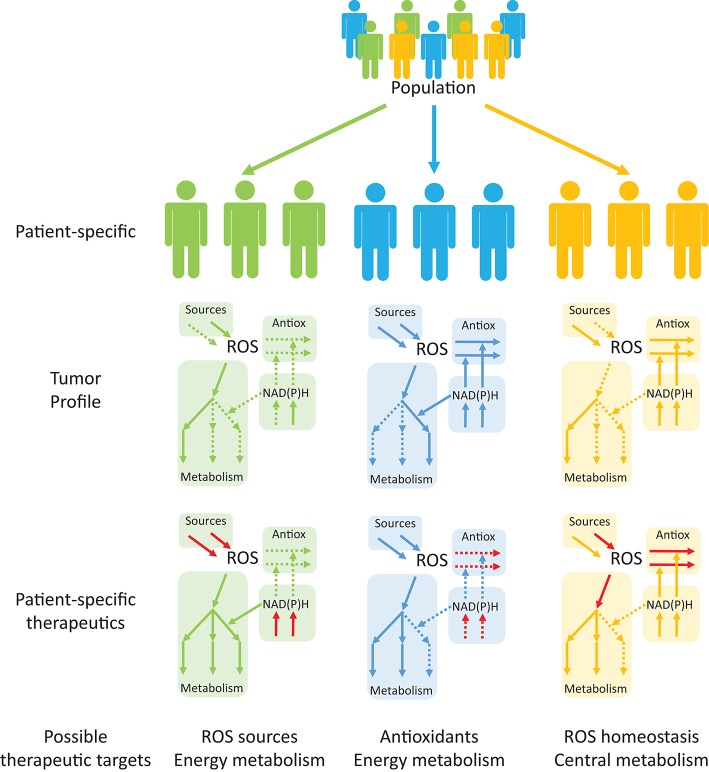
Reactive oxygen species (ROS) are important pathophysiological molecules involved in vital cellular processes. They are extremely harmful at high concentrations because they promote the generation of radicals and the oxidation of lipids, proteins, and nucleic acids, which can result in apoptosis. An imbalance of ROS and a disturbance of redox homeostasis are now recognized as a hallmark of complex diseases. Considering that ROS levels are significantly increased in cancer cells due to mitochondrial dysfunction, ROS metabolism has been targeted for the development of efficient treatment strategies, and antioxidants are used as potential chemotherapeutic drugs. However, initial ROS-focused clinical trials in which antioxidants were supplemented to patients provided inconsistent results, i.e., improved treatment or increased malignancy. These different outcomes may result from the highly heterogeneous redox responses of tumors in different patients. Hence, population-based treatment strategies are unsuitable and patient-tailored therapeutic approaches are required for the effective treatment of patients. Moreover, due to the crosstalk between ROS, reducing equivalents [e.g., NAD(P)H] and central metabolism, which is heterogeneous in cancer, finding the best therapeutic target requires the consideration of system-wide approaches that are capable of capturing the complex alterations observed in all of the associated pathways. Systems biology and engineering approaches may be employed to overcome these challenges, together with tools developed in personalized medicine. However, ROS- and redox-based therapies have yet to be addressed by these methodologies in the context of disease treatment. Here, we review the role of ROS and their coupled redox partners in tumorigenesis. Specifically, we highlight some of the challenges in understanding the role of hydrogen peroxide (H2O2), one of the most important ROS in pathophysiology in the progression of cancer. We also discuss its interplay with antioxidant defenses, such as the coupled peroxiredoxin/thioredoxin and glutathione/glutathione peroxidase systems, and its reducing equivalent metabolism. Finally, we highlight the need for system-level and patient-tailored approaches to clarify the roles of these systems and identify therapeutic targets through the use of the tools developed in personalized medicine.
Keywords: cancer heterogeneity; personalized medicine; reactive oxygen species; redox biology; systems biology.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources