

Osteopontin is linked with AKT, FoxO1, and myostatin in skeletal muscle cells
- PMID: 28745831
- PMCID: PMC5690863
- DOI: 10.1002/mus.25752
Osteopontin is linked with AKT, FoxO1, and myostatin in skeletal muscle cells
Abstract
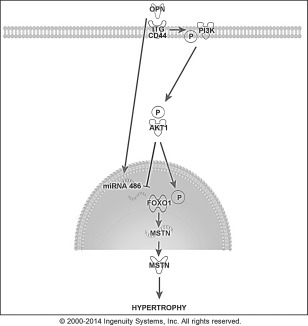
Introduction: Osteopontin (OPN) polymorphisms are associated with muscle size and modify disease progression in Duchenne muscular dystrophy (DMD). We hypothesized that OPN may share a molecular network with myostatin (MSTN).
Methods: Studies were conducted in the golden retriever (GRMD) and mdx mouse models of DMD. Follow-up in-vitro studies were employed in myogenic cells and the mdx mouse treated with recombinant mouse (rm) or human (Hu) OPN protein.
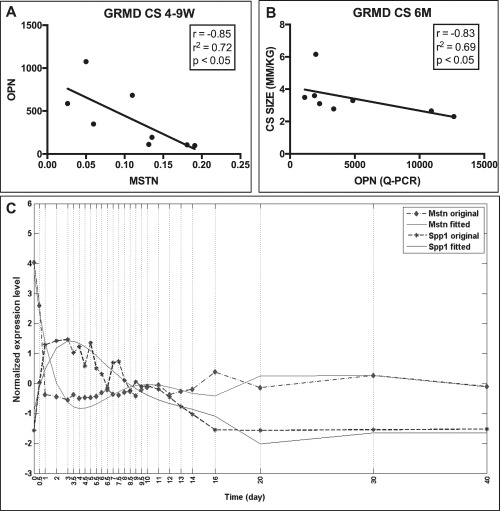
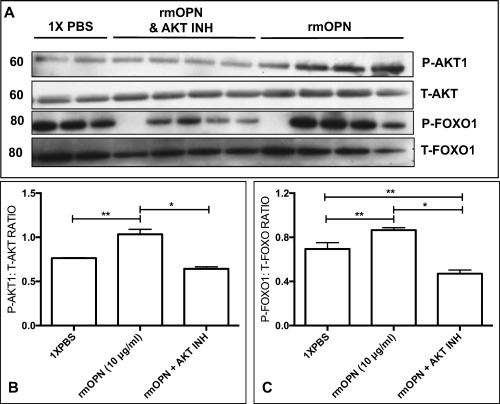
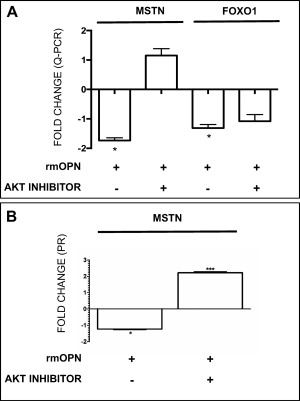
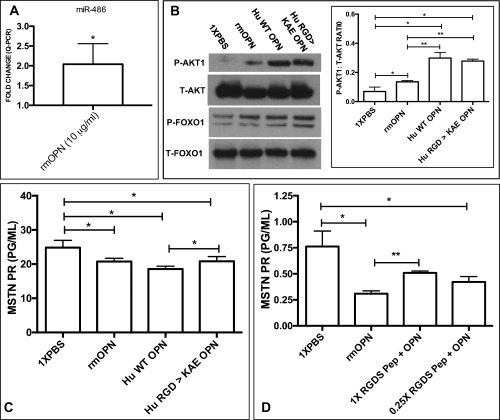
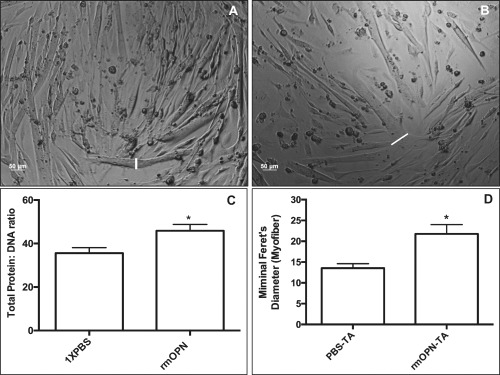
Results: OPN was increased and MSTN was decreased and levels correlated inversely in GRMD hypertrophied muscle. RM-OPN treatment led to induced AKT1 and FoxO1 phosphorylation, microRNA-486 modulation, and decreased MSTN. An AKT1 inhibitor blocked these effects, whereas an RGD-mutant OPN protein and an RGDS blocking peptide showed similar effects to the AKT inhibitor. RMOPN induced myotube hypertrophy and minimal Feret diameter in mdx muscle.
Discussion: OPN may interact with AKT1/MSTN/FoxO1 to modify normal and dystrophic muscle. Muscle Nerve 56: 1119-1127, 2017.
Keywords: AKT; Duchenne; GRMD; dog; mdx; muscle; myostatin; osteopontin.
© 2017 The Authors. Muscle & Nerve Published by Wiley Periodicals, Inc.
Figures

References
-
- Scatena M, Liaw L, Giachelli CM. Osteopontin: a multifunctional molecule regulating chronic inflammation and vascular disease. Arterioscler Thromb Vasc Biol 2007;27:2302–2309. - PubMed
-
- Liaw L, Almeida M, Hart CE, Schwartz SM, Giachelli CM. Osteopontin promotes vascular cell adhesion and spreading and is chemotactic for smooth muscle cells in vitro. Circ Res 1994;74:214–224. - PubMed
-
- Zheng DQ, Woodard AS, Tallini G, Languino LR. Substrate specificity of alpha(v)beta(3) integrin‐mediated cell migration and phosphatidylinositol 3‐kinase/AKT pathway activation. J Biol Chem 2000;275:24565–24574. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
