

Sequence Exchange between Homologous NB-LRR Genes Converts Virus Resistance into Nematode Resistance, and Vice Versa
- PMID: 28747428
- PMCID: PMC5580749
- DOI: 10.1104/pp.17.00485
Sequence Exchange between Homologous NB-LRR Genes Converts Virus Resistance into Nematode Resistance, and Vice Versa
Abstract
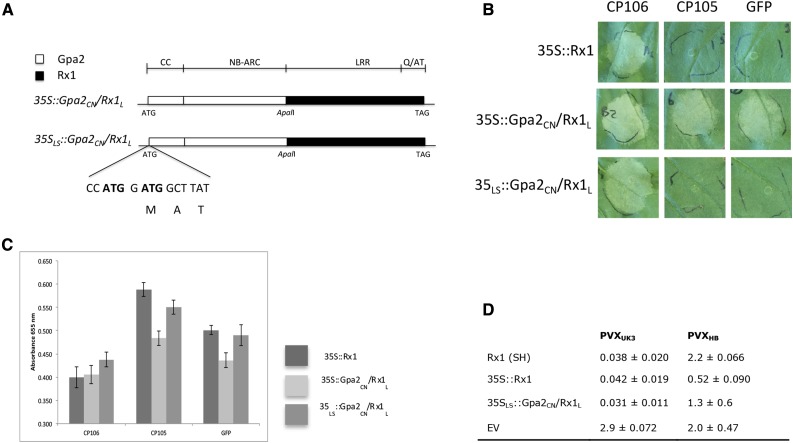
Plants have evolved a limited repertoire of NB-LRR disease resistance (R) genes to protect themselves against myriad pathogens. This limitation is thought to be counterbalanced by the rapid evolution of NB-LRR proteins, as only a few sequence changes have been shown to be sufficient to alter resistance specificities toward novel strains of a pathogen. However, little is known about the flexibility of NB-LRR R genes to switch resistance specificities between phylogenetically unrelated pathogens. To investigate this, we created domain swaps between the close homologs Gpa2 and Rx1, which confer resistance in potato (Solanum tuberosum) to the cyst nematode Globodera pallida and Potato virus X, respectively. The genetic fusion of the CC-NB-ARC of Gpa2 with the LRR of Rx1 (Gpa2CN/Rx1L) results in autoactivity, but lowering the protein levels restored its specific activation response, including extreme resistance to Potato virus X in potato shoots. The reciprocal chimera (Rx1CN/Gpa2L) shows a loss-of-function phenotype, but exchange of the first three LRRs of Gpa2 by the corresponding region of Rx1 was sufficient to regain a wild-type resistance response to G. pallida in the roots. These data demonstrate that exchanging the recognition moiety in the LRR is sufficient to convert extreme virus resistance in the leaves into mild nematode resistance in the roots, and vice versa. In addition, we show that the CC-NB-ARC can operate independently of the recognition specificities defined by the LRR domain, either aboveground or belowground. These data show the versatility of NB-LRR genes to generate resistance to unrelated pathogens with completely different lifestyles and routes of invasion.
© 2017 American Society of Plant Biologists. All Rights Reserved.
Figures



References
-
- Asai S, Shirasu K (2015) Plant cells under siege: plant immune system versus pathogen effectors. Curr Opin Plant Biol 28: 1–8 - PubMed
-
- Bakker E, Butterbach P, Rouppe van der Voort J, van der Vossen E, van Vliet J, Bakker J, Goverse A (2003) Genetic and physical mapping of homologues of the virus resistance gene Rx1 and the cyst nematode resistance gene Gpa2 in potato. Theor Appl Genet 106: 1524–1531 - PubMed
-
- Ballvora A, Jöcker A, Viehöver P, Ishihara H, Paal J, Meksem K, Bruggmann R, Schoof H, Weisshaar B, Gebhardt C (2007) Comparative sequence analysis of Solanum and Arabidopsis in a hot spot for pathogen resistance on potato chromosome V reveals a patchwork of conserved and rapidly evolving genome segments. BMC Genomics 8: 112. - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
