

Mutations in the E2 Glycoprotein and the 3' Untranslated Region Enhance Chikungunya Virus Virulence in Mice
- PMID: 28747508
- PMCID: PMC5625495
- DOI: 10.1128/JVI.00816-17
Mutations in the E2 Glycoprotein and the 3' Untranslated Region Enhance Chikungunya Virus Virulence in Mice
Abstract
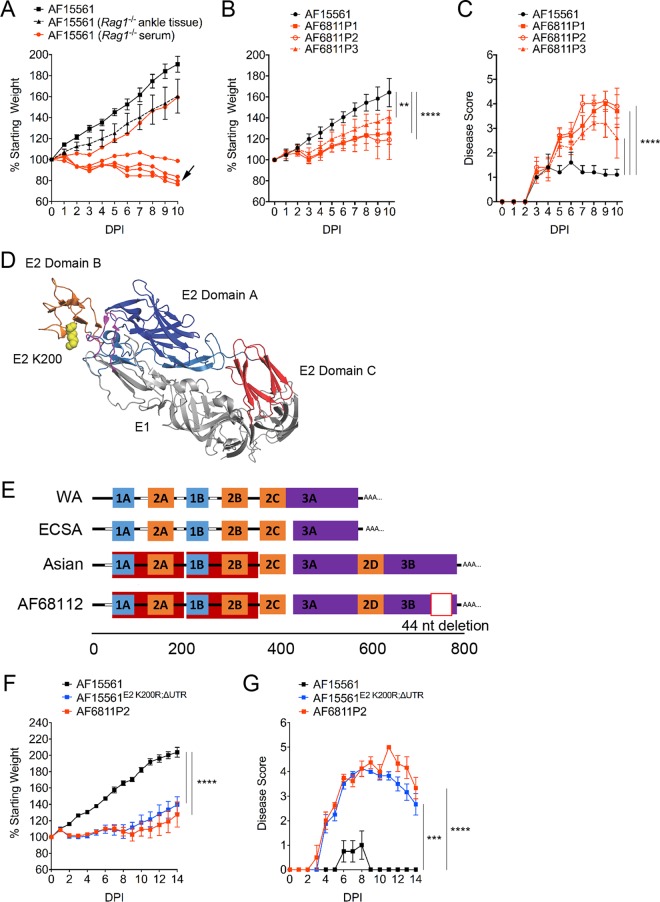
Chikungunya virus (CHIKV) is a mosquito-transmitted alphavirus that causes debilitating musculoskeletal pain and inflammation and can persist for months to years after acute infection. Although studies of humans and experimentally infected animals suggest that CHIKV infection persists in musculoskeletal tissues, the mechanisms for this remain poorly understood. To evaluate this further, we isolated CHIKV from the serum of persistently infected Rag1 -/- mice at day 28. When inoculated into naive wild-type (WT) mice, this persistently circulating CHIKV strain displayed a capacity for earlier dissemination and greater pathogenicity than the parental virus. Sequence analysis revealed a nonsynonymous mutation in the E2 glycoprotein (E2 K200R) and a deletion within the 3' untranslated region (3'-UTR). The introduction of these changes into the parental virus conferred enhanced virulence in mice, although primary tropism for musculoskeletal tissues was maintained. The E2 K200R mutation was largely responsible for enhanced viral dissemination and pathogenicity, although these effects were augmented by the 3'-UTR deletion. Finally, studies with Irf3/Irf7 -/- and Ifnar1 -/- mice suggest that the E2 K200R mutation enhances viral dissemination from the site of inoculation independently of interferon regulatory factor 3 (IRF3)-, IRF7-, and IFNAR1-mediated responses. As our findings reveal viral determinants of CHIKV dissemination and pathogenicity, their further study should help to elucidate host-virus interactions that determine acute and chronic CHIKV infection.IMPORTANCE CHIKV is a globally spreading, mosquito-transmitted virus that causes debilitating acute and chronic musculoskeletal disease in humans. The viral genetic determinants that dictate the severity of acute and chronic diseases are not understood. To improve our understanding of CHIKV pathogenesis, we evaluated a CHIKV strain isolated from the serum of chronically infected immunocompromised mice. Sequence analysis of this persistent CHIKV strain identified two mutations, an amino acid change in the E2 viral attachment protein and a deletion within the 3'-UTR of the viral genome. We identified roles for these mutations in the enhancement of viral dissemination from the inoculation site and in disease severity. These data improve our understanding of the viral determinants of CHIKV pathogenesis and adaptive changes that occur during viral persistence.
Keywords: alphavirus; chikungunya virus; viral pathogenesis; virulence determinants.
Copyright © 2017 American Society for Microbiology.
Figures
References
-
- Pan American Health Organization. 2015. Number of reported cases of chikungunya fever in the Americas, by country or territory. Pan American Health Organization, Washington, DC: http://www.paho.org/hq/index.php?option=com_topics&view=article&id=343& Accessed 15 May 2017.
-
- Economopoulou A, Dominguez M, Helynck B, Sissoko D, Wichmann O, Quenel P, Germonneau P, Quatresous I. 2009. Atypical Chikungunya virus infections: clinical manifestations, mortality and risk factors for severe disease during the 2005-2006 outbreak on Reunion. Epidemiol Infect 137:534–541. doi:10.1017/S0950268808001167. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
