

Mycobacteria Modify Their Cell Size Control under Sub-Optimal Carbon Sources
- PMID: 28748182
- PMCID: PMC5506092
- DOI: 10.3389/fcell.2017.00064
Mycobacteria Modify Their Cell Size Control under Sub-Optimal Carbon Sources
Abstract
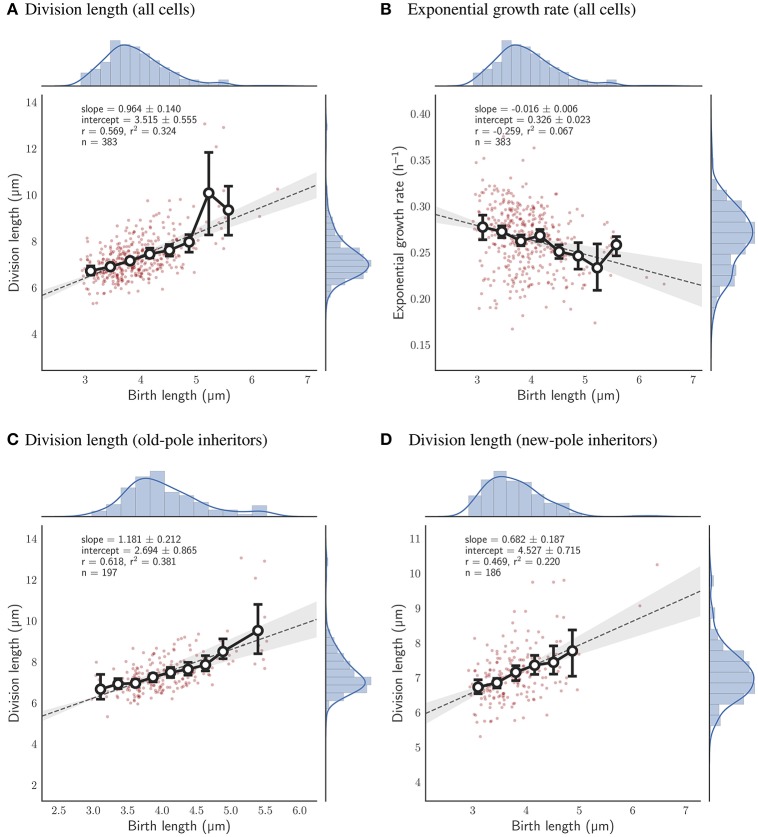
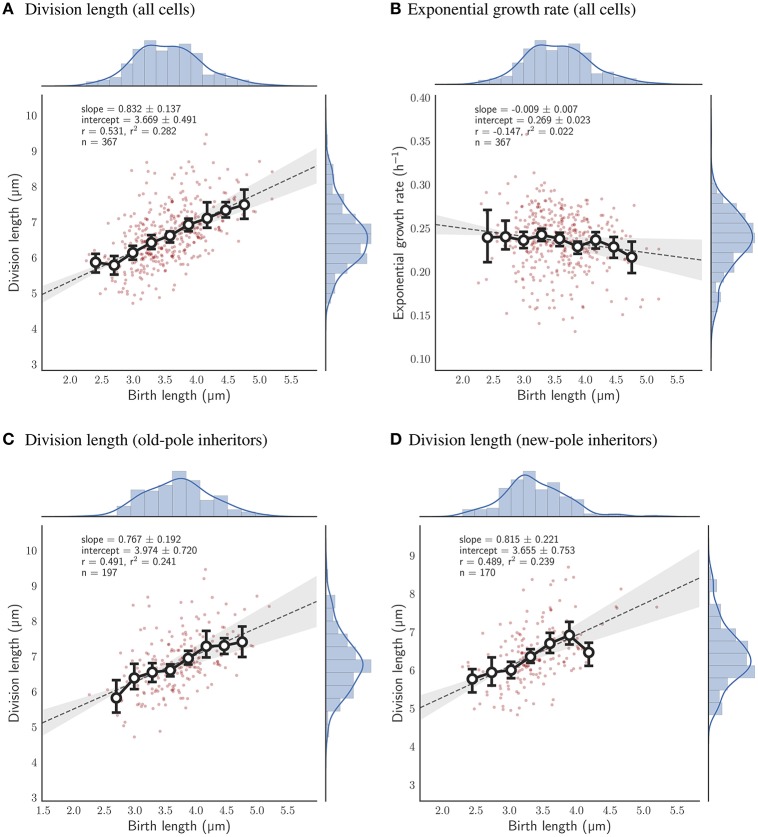
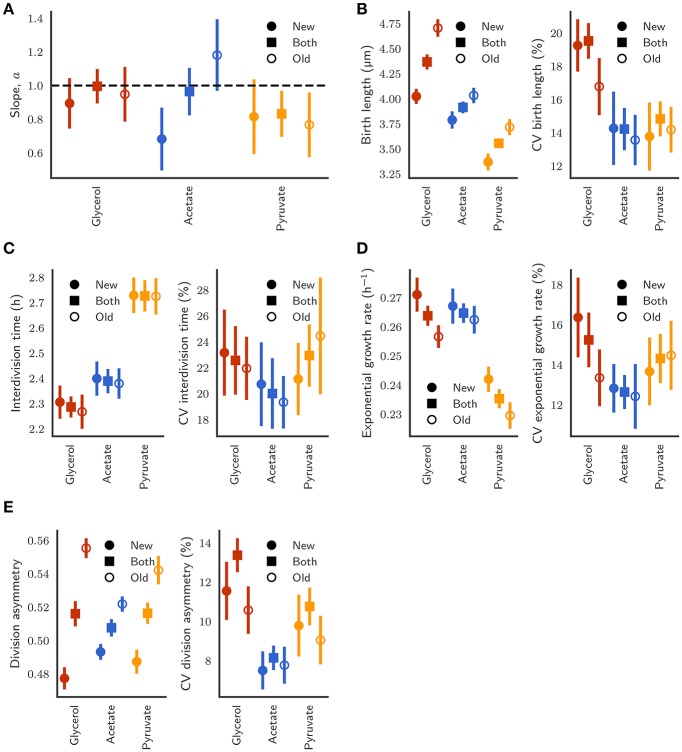
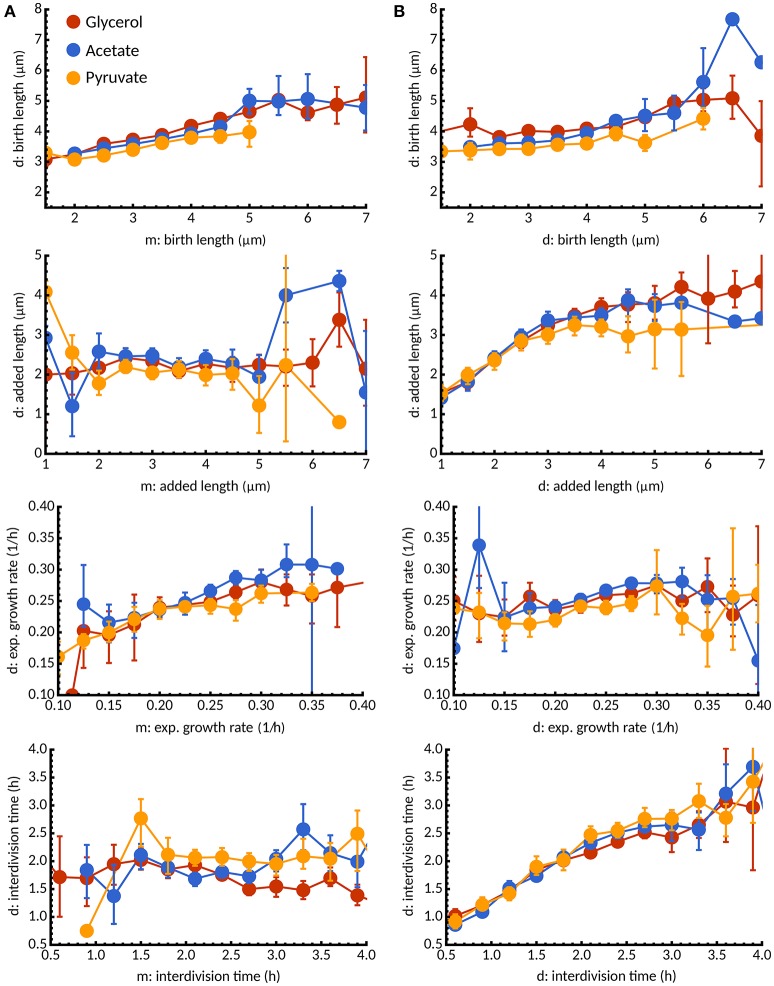
The decision to divide is the most important one that any cell must make. Recent single cell studies suggest that most bacteria follow an "adder" model of cell size control, incorporating a fixed amount of cell wall material before dividing. Mycobacteria, including the causative agent of tuberculosis Mycobacterium tuberculosis, are known to divide asymmetrically resulting in heterogeneity in growth rate, doubling time, and other growth characteristics in daughter cells. The interplay between asymmetric cell division and adder size control has not been extensively investigated. Moreover, the impact of changes in the environment on growth rate and cell size control have not been addressed for mycobacteria. Here, we utilize time-lapse microscopy coupled with microfluidics to track live Mycobacterium smegmatis cells as they grow and divide over multiple generations, under a variety of growth conditions. We demonstrate that, under optimal conditions, M. smegmatis cells robustly follow the adder principle, with constant added length per generation independent of birth size, growth rate, and inherited pole age. However, the nature of the carbon source induces deviations from the adder model in a manner that is dependent on pole age. Understanding how mycobacteria maintain cell size homoeostasis may provide crucial targets for the development of drugs for the treatment of tuberculosis, which remains a leading cause of global mortality.
Keywords: adder; asymmetric cell division; cell size control; mycobacteria; sizer.
Figures
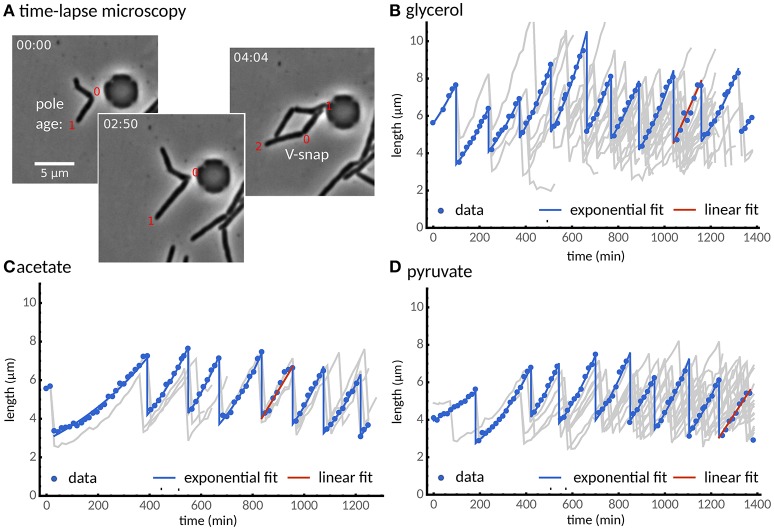
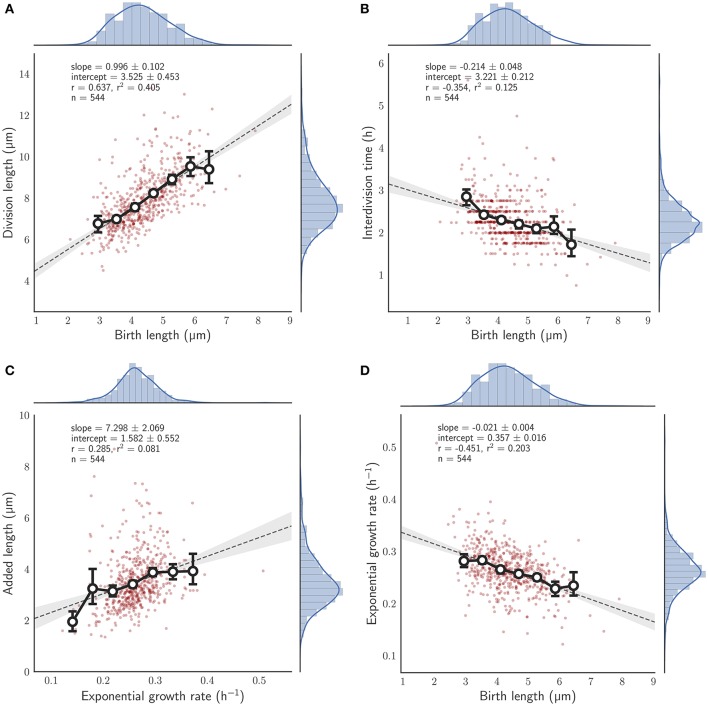
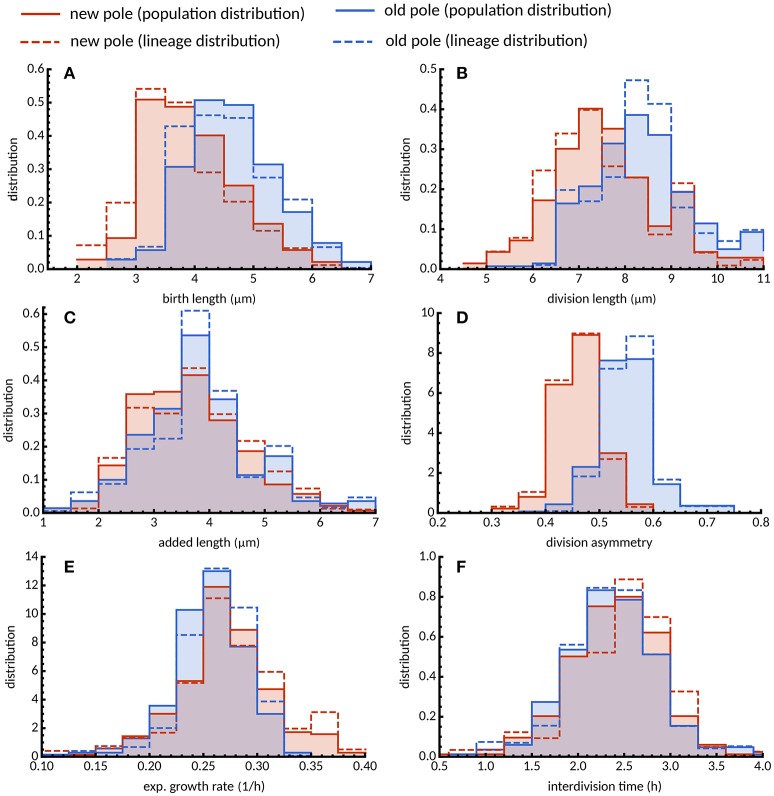
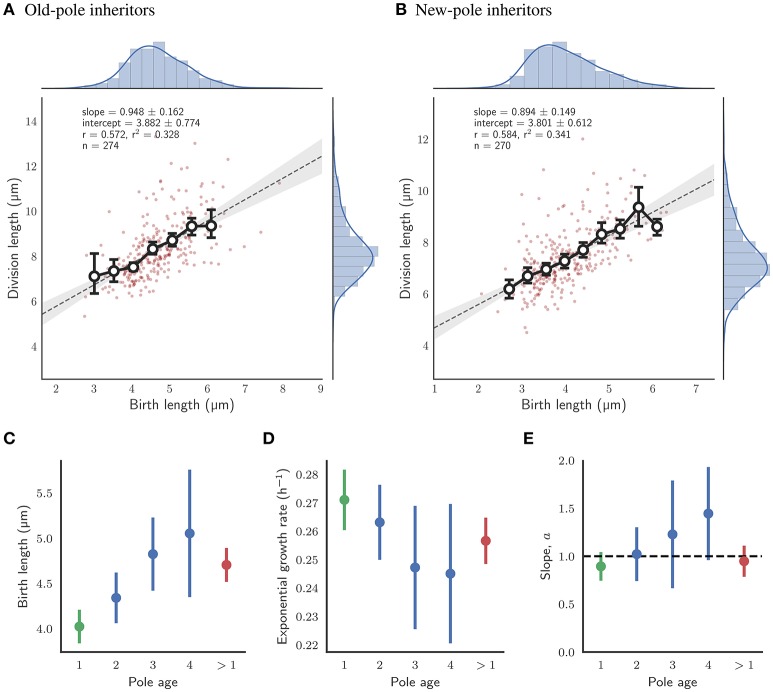
References
-
- Amir A. (2014). Cell size regulation in bacteria. Phys. Rev. Lett. 112:208102 10.1103/physrevlett.112.208102 - DOI
-
- Bertaux F., Von Kügelgen J., Marguerat S., Shahrezaei V. (2016). A unified coarse-grained theory of bacterial physiology explains the relationship between cell size, growth rate and proteome composition under various growth limitations. bioRxiv 078998. 10.1101/078998 - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources