

Mechanisms of protein homeostasis (proteostasis) maintain stem cell identity in mammalian pluripotent stem cells
- PMID: 28748323
- PMCID: PMC11105389
- DOI: 10.1007/s00018-017-2602-1
Mechanisms of protein homeostasis (proteostasis) maintain stem cell identity in mammalian pluripotent stem cells
Abstract
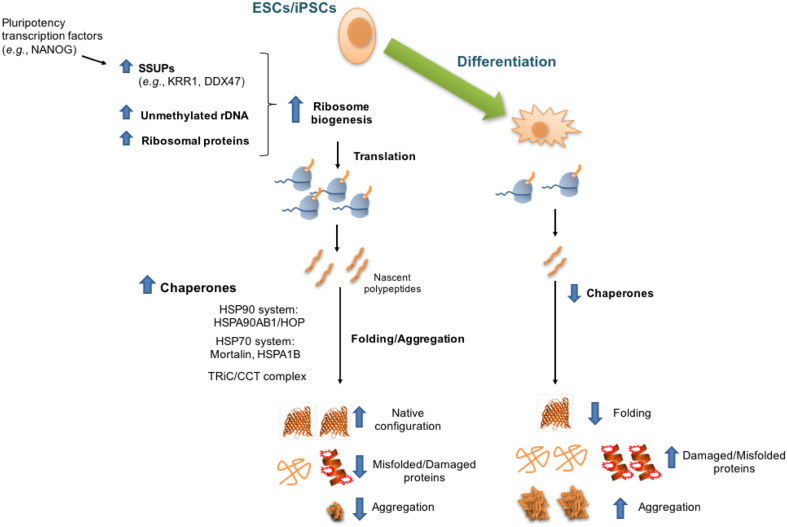
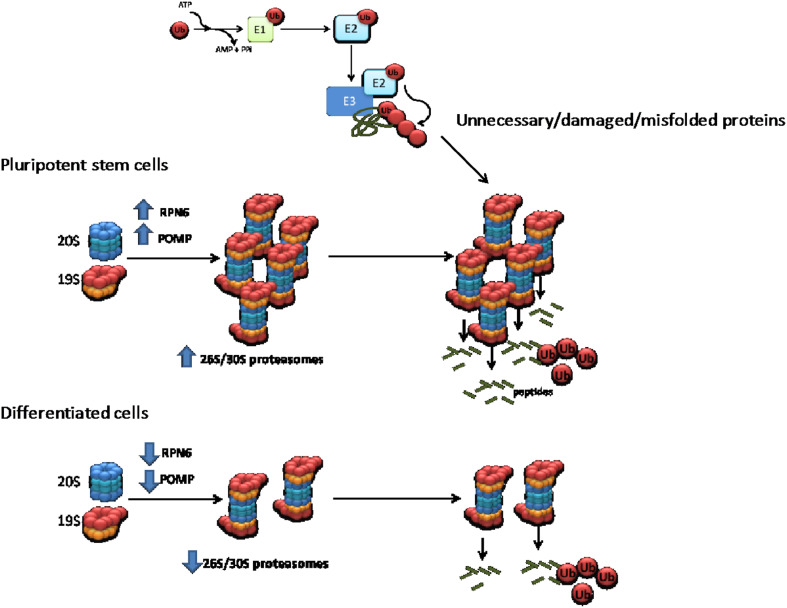
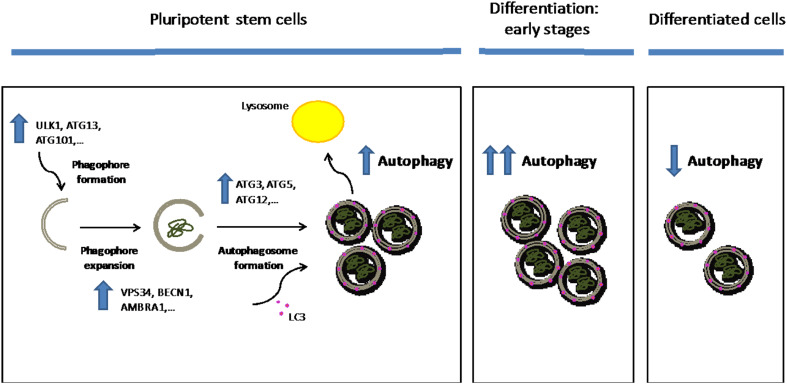
Protein homeostasis, or proteostasis, is essential for cell function, development, and organismal viability. The composition of the proteome is adjusted to the specific requirements of a particular cell type and status. Moreover, multiple metabolic and environmental conditions challenge the integrity of the proteome. To maintain the quality of the proteome, the proteostasis network monitors proteins from their synthesis through their degradation. Whereas somatic stem cells lose their ability to maintain proteostasis with age, immortal pluripotent stem cells exhibit a stringent proteostasis network associated with their biological function and intrinsic characteristics. Moreover, growing evidence indicates that enhanced proteostasis mechanisms play a central role in immortality and cell fate decisions of pluripotent stem cells. Here, we will review new insights into the melding fields of proteostasis and pluripotency and their implications for the understanding of organismal development and survival.
Keywords: Autophagy; Chaperones; Differentiation; Pluripotency; Proteasome; Proteostasis; Stress responses.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
