

Cross-modulation of pathogen-specific pathways enhances malnutrition during enteric co-infection with Giardia lamblia and enteroaggregative Escherichia coli
- PMID: 28750066
- PMCID: PMC5549954
- DOI: 10.1371/journal.ppat.1006471
Cross-modulation of pathogen-specific pathways enhances malnutrition during enteric co-infection with Giardia lamblia and enteroaggregative Escherichia coli
Abstract
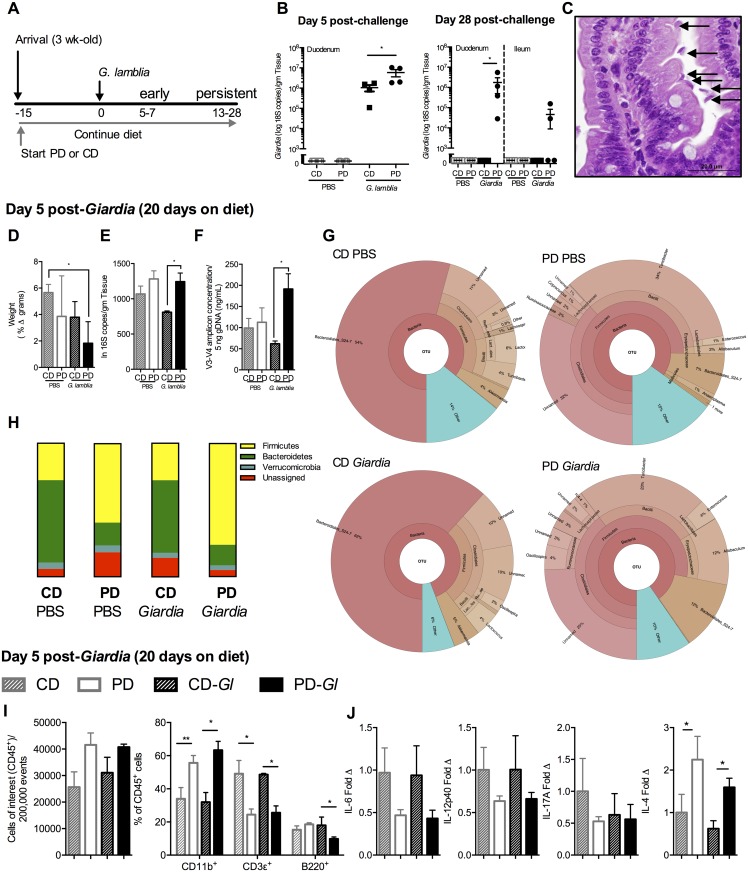
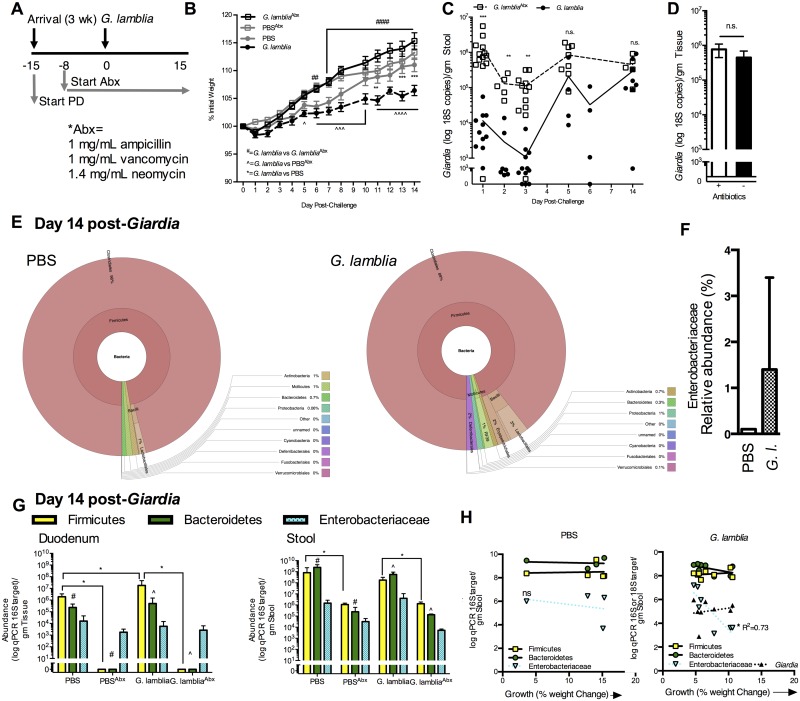
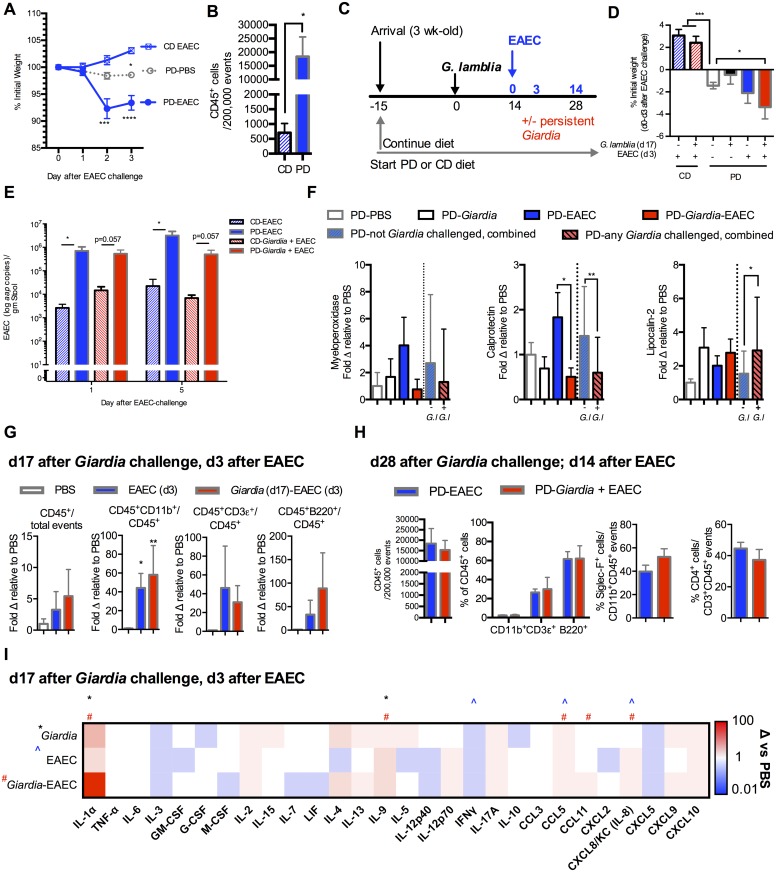
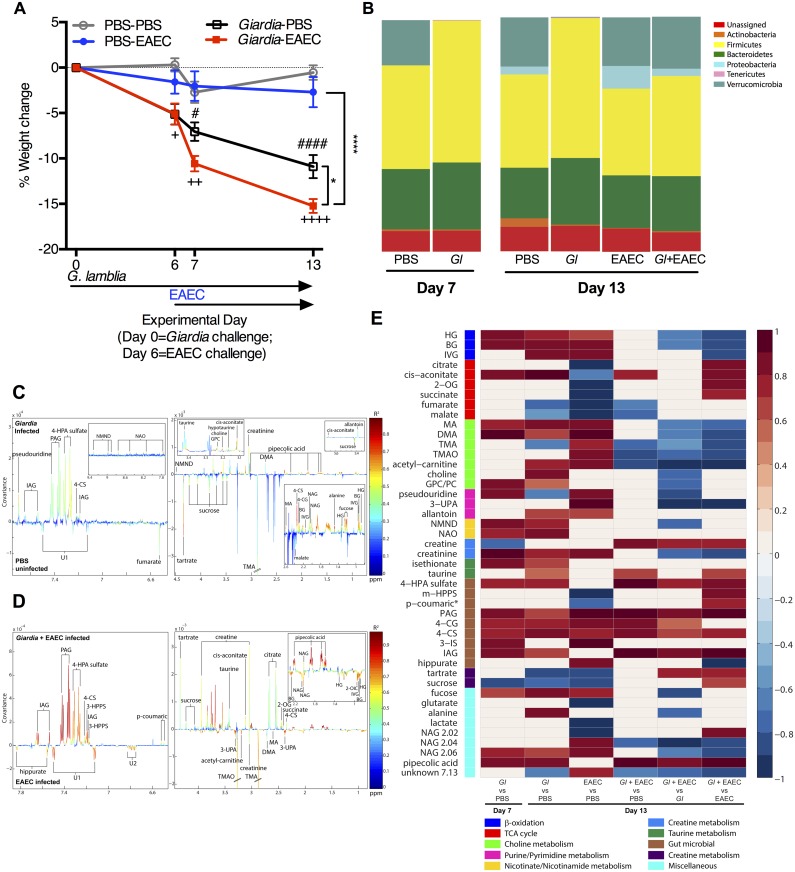
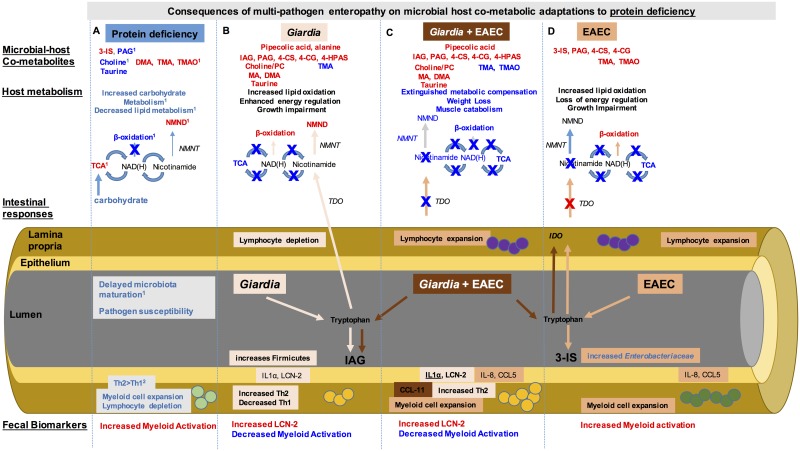
Diverse enteropathogen exposures associate with childhood malnutrition. To elucidate mechanistic pathways whereby enteric microbes interact during malnutrition, we used protein deficiency in mice to develop a new model of co-enteropathogen enteropathy. Focusing on common enteropathogens in malnourished children, Giardia lamblia and enteroaggregative Escherichia coli (EAEC), we provide new insights into intersecting pathogen-specific mechanisms that enhance malnutrition. We show for the first time that during protein malnutrition, the intestinal microbiota permits persistent Giardia colonization and simultaneously contributes to growth impairment. Despite signals of intestinal injury, such as IL1α, Giardia-infected mice lack pro-inflammatory intestinal responses, similar to endemic pediatric Giardia infections. Rather, Giardia perturbs microbial host co-metabolites of proteolysis during growth impairment, whereas host nicotinamide utilization adaptations that correspond with growth recovery increase. EAEC promotes intestinal inflammation and markers of myeloid cell activation. During co-infection, intestinal inflammatory signaling and cellular recruitment responses to EAEC are preserved together with a Giardia-mediated diminishment in myeloid cell activation. Conversely, EAEC extinguishes markers of host energy expenditure regulatory responses to Giardia, as host metabolic adaptations appear exhausted. Integrating immunologic and metabolic profiles during co-pathogen infection and malnutrition, we develop a working mechanistic model of how cumulative diet-induced and pathogen-triggered microbial perturbations result in an increasingly wasted host.
Conflict of interest statement
I have read the journal's policy and the authors of this manuscript have the following competing interests: LAB was a temporary consultant for Lupin Pharmaceuticals, June - December 2015.
Figures
Comment in
-
Infections: Feeding the microbial multitudes: co-infection in a malnourished host.Nat Rev Gastroenterol Hepatol. 2017 Dec;14(12):695-696. doi: 10.1038/nrgastro.2017.133. Epub 2017 Sep 27. Nat Rev Gastroenterol Hepatol. 2017. PMID: 28951583 No abstract available.
References
-
- Smith MI, Yatsunenko T, Manary MJ, Trehan I, Mkakosya R, Cheng J, et al. Gut microbiomes of Malawian twin pairs discordant for kwashiorkor. Science. 2013;339(6119):548–54. Epub 2013/02/01. doi: 10.1126/science.1229000 ; - DOI - PMC - PubMed
-
- Blanton LV, Charbonneau MR, Salih T, Barratt MJ, Venkatesh S, Ilkaveya O, et al. Gut bacteria that prevent growth impairments transmitted by microbiota from malnourished children. Science. 2016;351(6275). doi: 10.1126/science.aad3311 ; - DOI - PMC - PubMed
-
- Guerrant RL, Leite AM, Pinkerton R, Medeiros PH, Cavalcante PA, DeBoer M, et al. Biomarkers of Environmental Enteropathy, Inflammation, Stunting, and Impaired Growth in Children in Northeast Brazil. PLoS One. 2016;11(9):e0158772 doi: 10.1371/journal.pone.0158772 ; - DOI - PMC - PubMed
-
- Taniuchi M, Platts-Mills JA, Begum S, Uddin MJ, Sobuz SU, Liu J, et al. Impact of enterovirus and other enteric pathogens on oral polio and rotavirus vaccine performance in Bangladeshi infants. Vaccine. 2016;34(27):3068–75. Epub 2016/05/08. doi: 10.1016/j.vaccine.2016.04.080 ; - DOI - PMC - PubMed
-
- Prata MMG H A, Bolick DT, Pinkerton R, Lima AMM, Guerrant RL. Comparisons between myeloperoxidase, lactoferrin, calprotectin and lipocalin-2, as fecal biomarkers of intestinal inflammation in malnourished children. J Transl Sci. 2016;2(2):134–9. doi: 10.15761/JTS.1000130 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
