

SID-1 Functions in Multiple Roles To Support Parental RNAi in Caenorhabditis elegans
- PMID: 28751423
- PMCID: PMC5629322
- DOI: 10.1534/genetics.117.300067
SID-1 Functions in Multiple Roles To Support Parental RNAi in Caenorhabditis elegans
Abstract
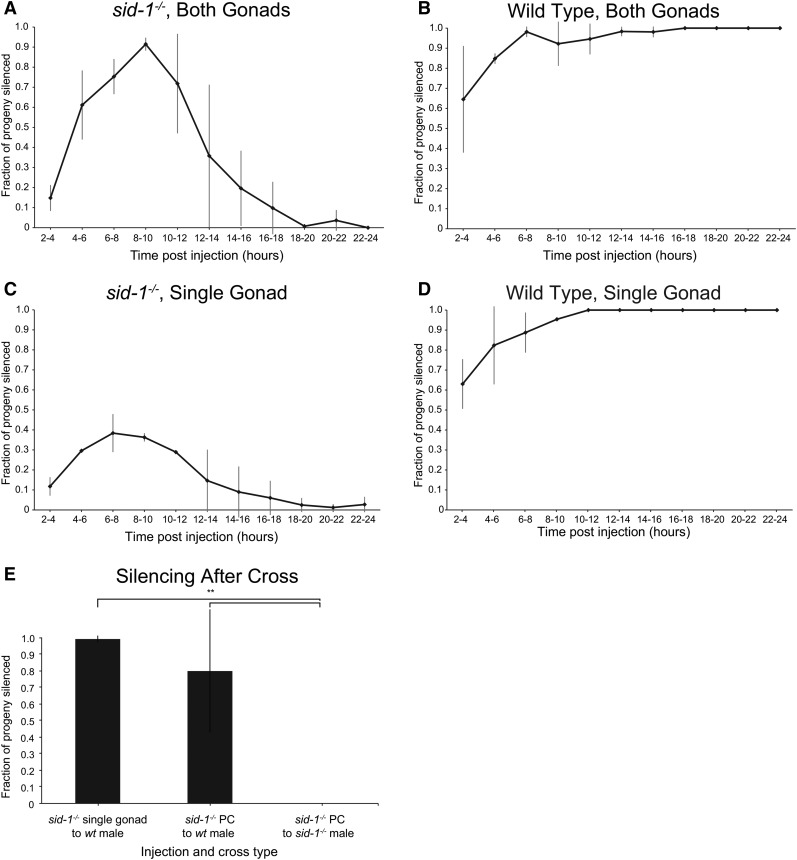
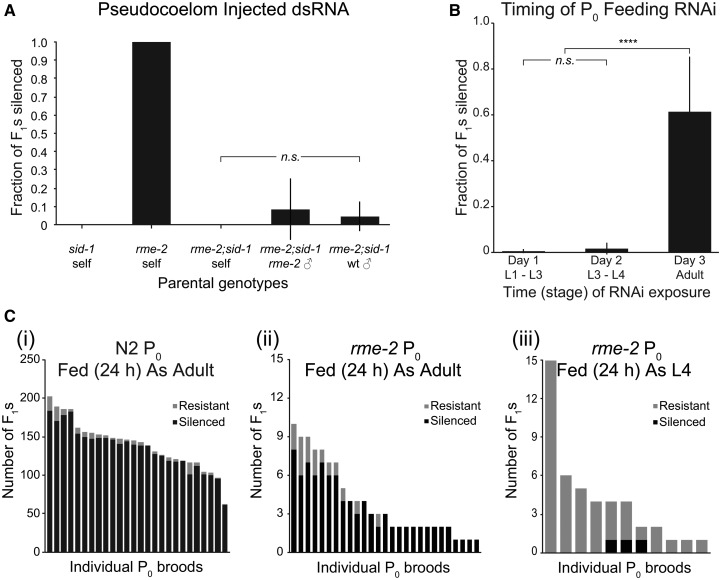
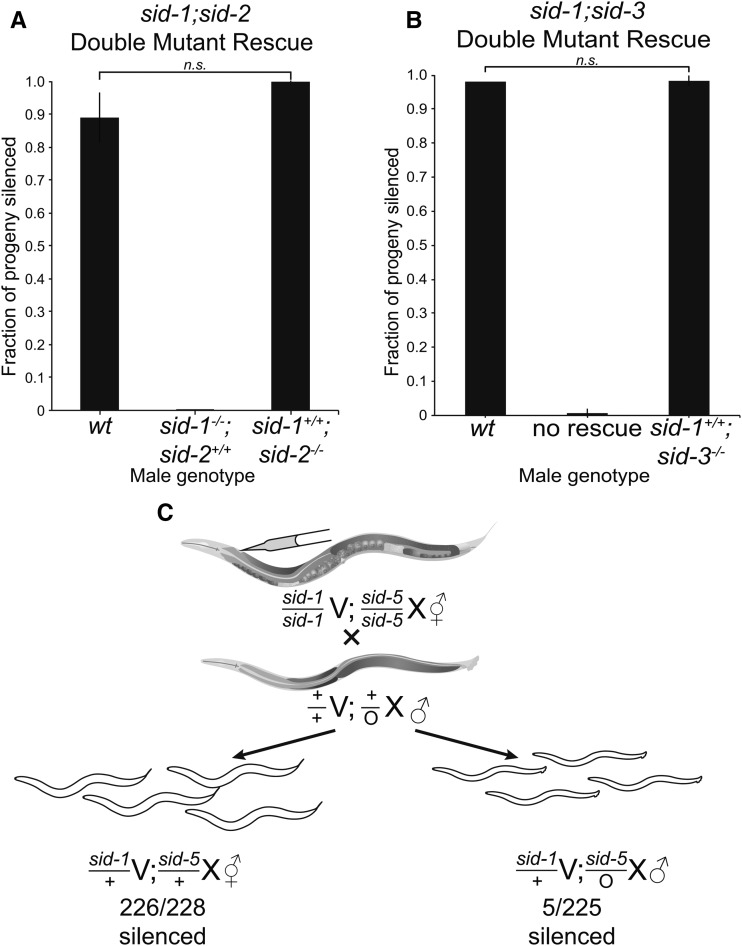
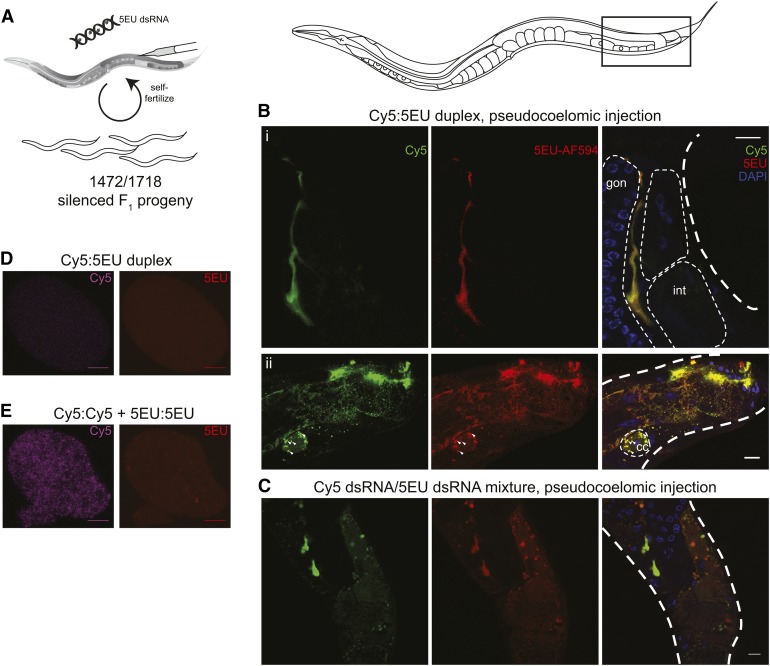
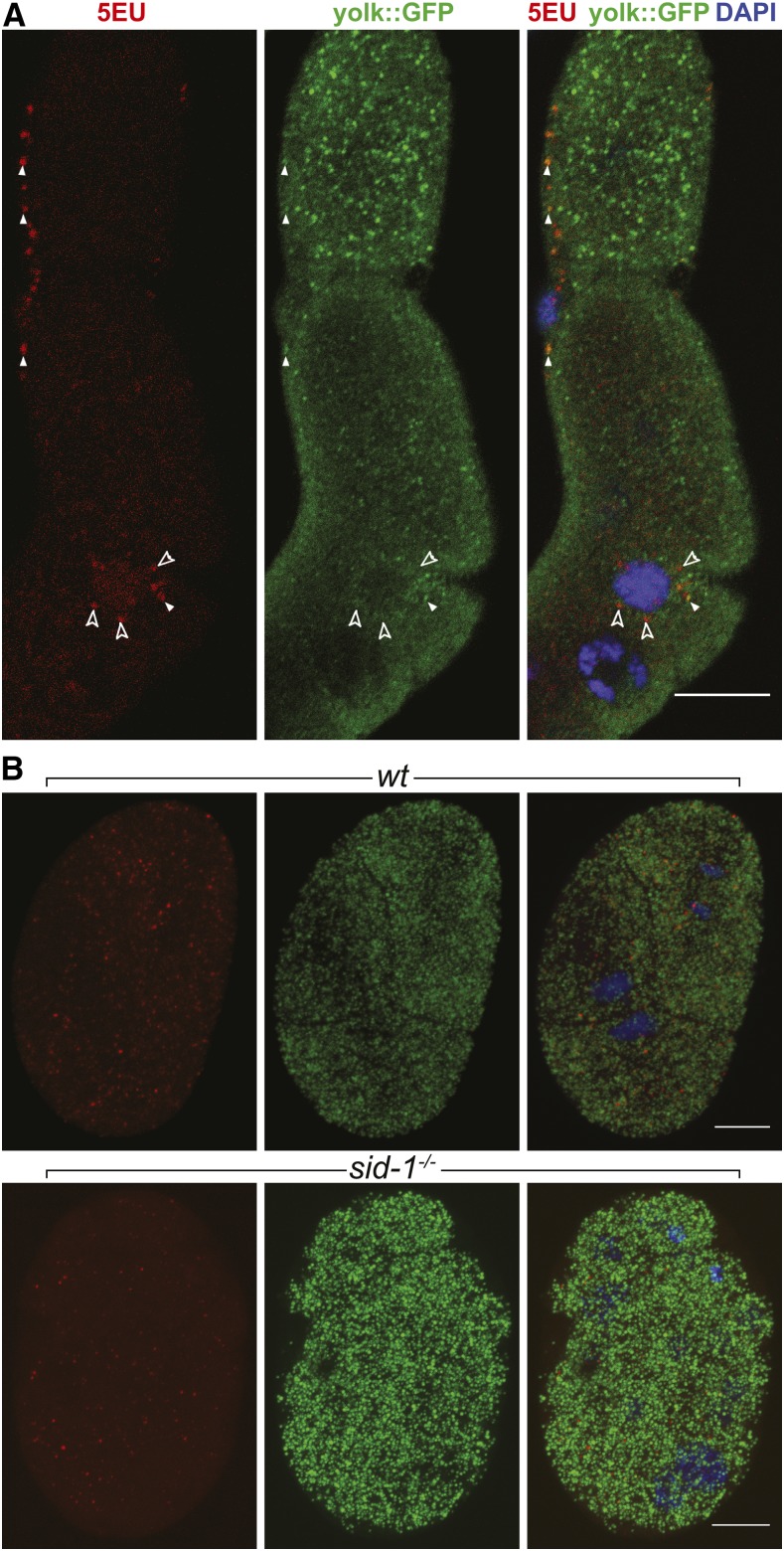
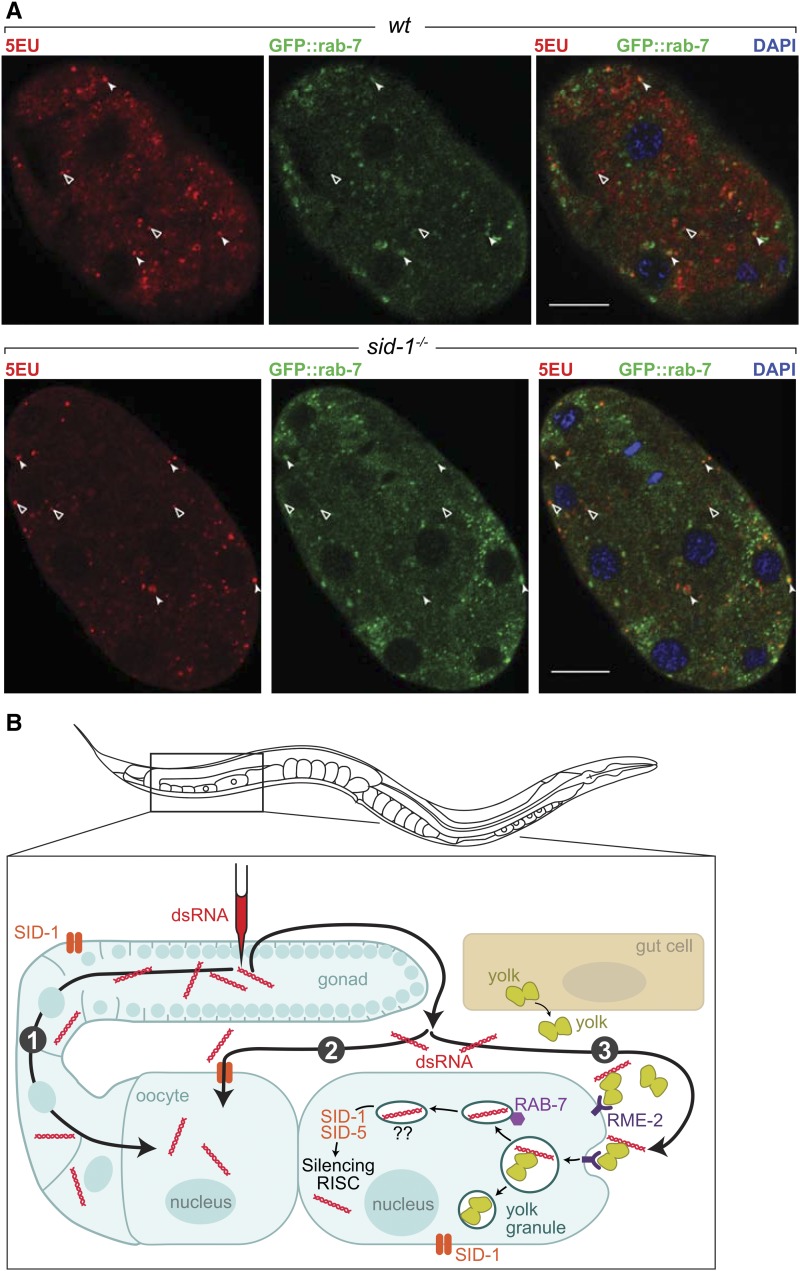
Systemic RNA interference (RNAi) in Caenorhbaditis elegans requires sid-1, sid-3, and sid-5 Injected, expressed, or ingested double-stranded RNA (dsRNA) is transported between cells, enabling RNAi in most tissues, including the germline and progeny (parental RNAi). A recent report claims that parental RNAi also requires the yolk receptor rme-2 Here, we characterize the role of the sid genes and rme-2 in parental RNAi. We identify multiple independent paths for maternal dsRNA to reach embryos and initiate RNAi. We showed previously that maternal and embryonic sid-1 contribute independently to parental RNAi. Here we demonstrate a role for embryonic sid-5, but not sid-2 or sid-3 in parental RNAi. We also find that maternal rme-2 contributes to but is not required for parental RNAi. We determine that parental RNAi by feeding occurs nearly exclusively in adults. We also introduce 5-ethynyluridine to densely internally label dsRNA, avoiding complications associated with other labeling strategies such as inhibition of normal dsRNA trafficking and separation of label and RNA. Labeling shows that yolk and dsRNA do not colocalize following endocytosis, suggesting independent uptake, and, furthermore, dsRNA appears to rapidly progress through the RAB-7 endocytosis pathway independently of sid-1 activity. Our results support the premise that although sid-1 functions in multiple roles, it alone is central and absolutely required for inheritance of silencing RNAs.
Keywords: Caenorhabditis elegans; RNAi; double-stranded RNA; yolk.
Copyright © 2017 by the Genetics Society of America.
Figures
References
-
- Cappelle K., de Oliveira C. F. R., Eynde V. B., Christiaens O., Smagghe G., 2016. The involvement of clathrin-mediated endocytosis and two Sid-1-like transmembrane proteins in double-stranded RNA uptake in the Colorado potato beetle midgut. Insect Mol. Biol. 25: 315–323. - PubMed
-
- Feinberg E. H., Hunter C. P., 2003. Transport of dsRNA into cells by the transmembrane protein SID-1. Science 301: 1545–1547. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
