

Control of species-dependent cortico-motoneuronal connections underlying manual dexterity
- PMID: 28751609
- PMCID: PMC5774341
- DOI: 10.1126/science.aan3721
Control of species-dependent cortico-motoneuronal connections underlying manual dexterity
Abstract
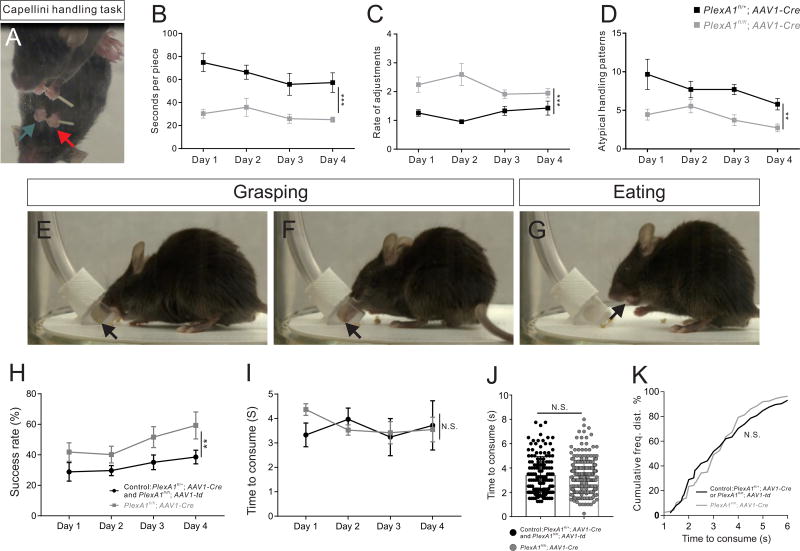
Superior manual dexterity in higher primates emerged together with the appearance of cortico-motoneuronal (CM) connections during the evolution of the mammalian corticospinal (CS) system. Previously thought to be specific to higher primates, we identified transient CM connections in early postnatal mice, which are eventually eliminated by Sema6D-PlexA1 signaling. PlexA1 mutant mice maintain CM connections into adulthood and exhibit superior manual dexterity as compared with that of controls. Last, differing PlexA1 expression in layer 5 of the motor cortex, which is strong in wild-type mice but weak in humans, may be explained by FEZF2-mediated cis-regulatory elements that are found only in higher primates. Thus, species-dependent regulation of PlexA1 expression may have been crucial in the evolution of mammalian CS systems that improved fine motor control in higher primates.
Copyright © 2017 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures



Comment in
-
Motor systems: Mice get manual.Nat Rev Neurosci. 2017 Aug 21;18(9):512. doi: 10.1038/nrn.2017.108. Nat Rev Neurosci. 2017. PMID: 28824179 No abstract available.
References
-
- Krubitzer L. The magnificent compromise: cortical field evolution in mammals. Neuron. 2007;56:201–208. - PubMed
-
- Lemon RN. Descending pathways in motor control. Annu Rev Neurosci. 2008;31:195–218. - PubMed
-
- Yang HW, Lemon RN. An electron microscopic examination of the corticospinal projection to the cervical spinal cord in the rat: lack of evidence for cortico-motoneuronal synapses. Exp Brain Res. 2003;149:458–469. - PubMed
-
- Alstermark B, Ogawa J. In vivo recordings of bulbospinal excitation in adult mouse forelimb motoneurons. J Neurophysiol. 2004;92:1958–1962. - PubMed
-
- Alstermark B, Ogawa J, Isa T. Lack of monosynaptic corticomotoneuronal EPSPs in rats: disynaptic EPSPs mediated via reticulospinal neurons and polysynaptic EPSPs via segmental interneurons. J Neurophysiol. 2004;91:1832–1839. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
