

Increased Ca buffering underpins remodelling of Ca2+ handling in old sheep atrial myocytes
- PMID: 28752958
- PMCID: PMC5621500
- DOI: 10.1113/JP274053
Increased Ca buffering underpins remodelling of Ca2+ handling in old sheep atrial myocytes
Abstract
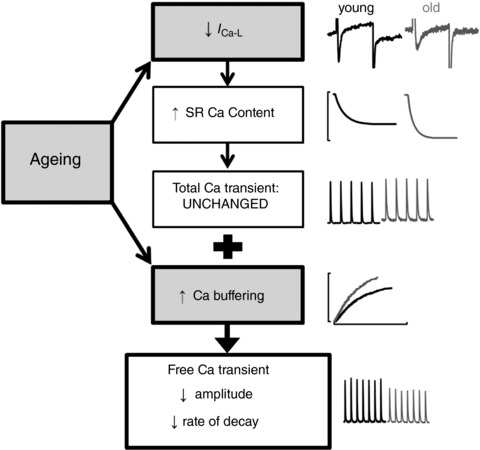
Key points: Ageing is associated with an increased risk of cardiovascular disease and arrhythmias, with the most common arrhythmia being found in the atria of the heart. Little is known about how the normal atria of the heart remodel with age and thus why dysfunction might occur. We report alterations to the atrial systolic Ca2+ transient that have implications for the function of the atrial in the elderly. We describe a novel mechanism by which increased Ca buffering can account for changes to systolic Ca2+ in the old atria. The present study helps us to understand how the processes regulating atrial contraction are remodelled during ageing and provides a basis for future work aiming to understand why dysfunction develops.
Abstract: Many cardiovascular diseases, including those affecting the atria, are associated with advancing age. Arrhythmias, including those in the atria, can arise as a result of electrical remodelling or alterations in Ca2+ homeostasis. In the atria, age-associated changes in the action potential have been documented. However, little is known about remodelling of intracellular Ca2+ homeostasis in the healthy aged atria. Using single atrial myocytes from young and old Welsh Mountain sheep, we show the free Ca2+ transient amplitude and rate of decay of systolic Ca2+ decrease with age, whereas sarcoplasmic reticulum (SR) Ca content increases. An increase in intracellular Ca buffering explains both the decrease in Ca2+ transient amplitude and decay kinetics in the absence of any change in sarcoendoplasmic reticulum calcium transport ATPase function. Ageing maintained the integrated Ca2+ influx via ICa-L but decreased peak ICa-L . Decreased peak ICa-L was found to be responsible for the age-associated increase in SR Ca content but not the decrease in Ca2+ transient amplitude. Instead, decreased peak ICa-L offsets increased SR load such that Ca2+ release from the SR was maintained during ageing. The results of the present study highlight a novel mechanism by which increased Ca buffering decreases systolic Ca2+ in old atria. Furthermore, for the first time, we have shown that SR Ca content is increased in old atrial myocytes.
Keywords: age; atria; buffering; calcium.
© 2017 The Authors. The Journal of Physiology published by John Wiley & Sons Ltd on behalf of The Physiological Society.
Figures






References
-
- Abhishekh HA, Nisarga P, Kisan R, Meghana A, Chandran S, Trichur R & Sathyaprabha TN (2013). Influence of age and gender on autonomic regulation of heart. J Clin Monit Comput 27, 259–264. - PubMed
-
- Anyukhovsky EP, Sosunov EA, Chandra P, Rosen TS, Boyden PA, Danilo P, Jr. & Rosen MR (2005). Age‐associated changes in electrophysiologic remodeling: a potential contributor to initiation of atrial fibrillation. Cardiovasc Res 66, 353–363. - PubMed
-
- Bers DM (2001). Excitation‐Contraction Coupling and Cardiac Contractile Force, (2 ed.) Kluwer Academic Publishers, Dordrecht.
-
- Boyd AC, Schiller NB, Leung D, Ross DL & Thomas L (2011). Atrial dilation and altered function are mediated by age and diastolic function but not before the eighth decade. JACC Cardiovasc Imaging 4, 234–242. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous