

The perilipin family of lipid droplet proteins: Gatekeepers of intracellular lipolysis
- PMID: 28754637
- PMCID: PMC5595658
- DOI: 10.1016/j.bbalip.2017.07.009
The perilipin family of lipid droplet proteins: Gatekeepers of intracellular lipolysis
Abstract
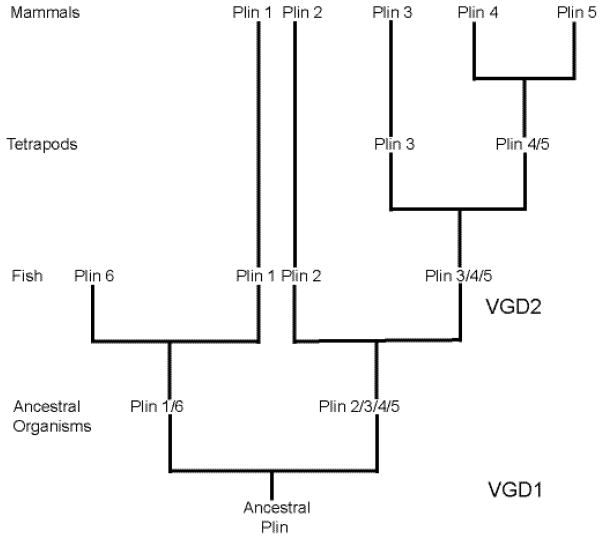
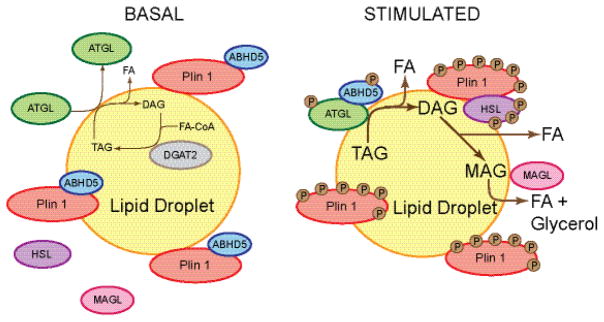
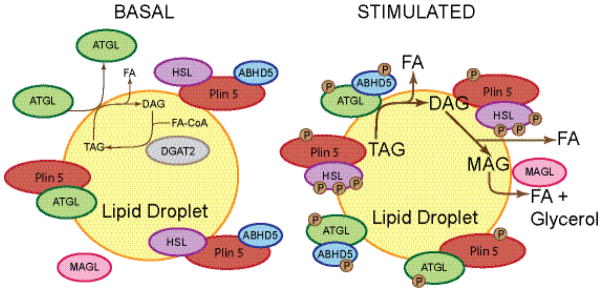
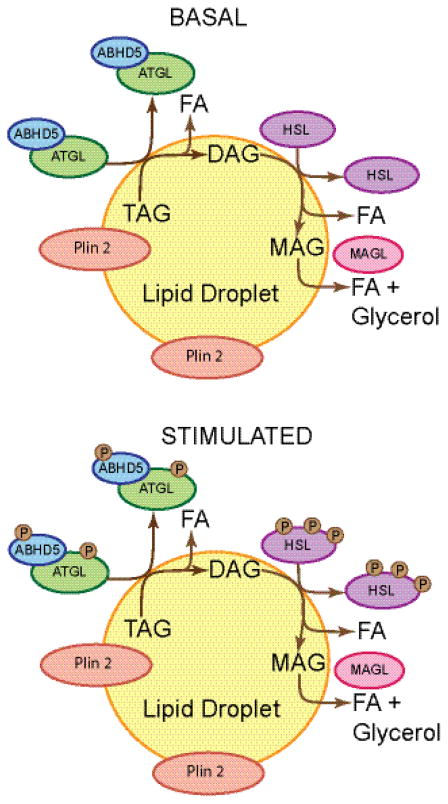
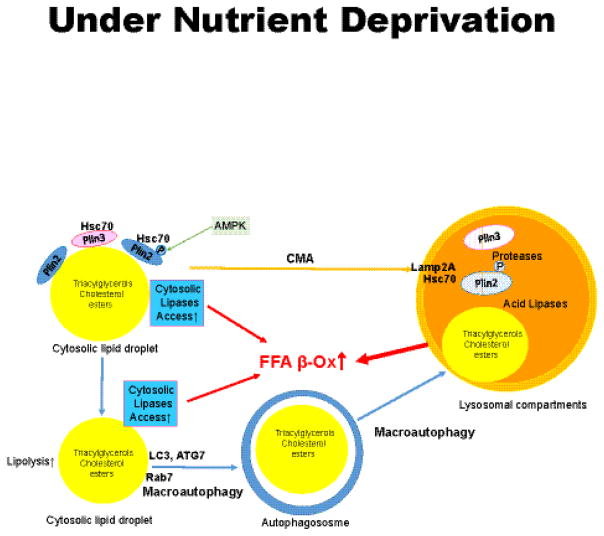
Lipid droplets in chordates are decorated by two or more members of the perilipin family of lipid droplet surface proteins. The perilipins sequester lipids by protecting lipid droplets from lipase action. Their relative expression and protective nature is adapted to the balance of lipid storage and utilization in specific cells. Most cells of the body have tiny lipid droplets with perilipins 2 and 3 at the surfaces, whereas specialized fat-storing cells with larger lipid droplets also express perilipins 1, 4, and/or 5. Perilipins 1, 2, and 5 modulate lipolysis by controlling the access of lipases and co-factors of lipases to substrate lipids stored within lipid droplets. Although perilipin 2 is relatively permissive to lipolysis, perilipins 1 and 5 have distinct control mechanisms that are altered by phosphorylation. Here we evaluate recent progress toward understanding functions of the perilipins with a focus on their role in regulating lipolysis and autophagy. This article is part of a Special Issue entitled: Recent Advances in Lipid Droplet Biology edited by Rosalind Coleman and Matthijs Hesselink.
Keywords: ABHD5; Adipose triglyceride lipase; Autophagy; Hormone-sensitive lipase; Lipid droplet; Lipolysis; Monoacylglycerol lipase; Perilipin; Triacylglycerol.
Copyright © 2017 Elsevier B.V. All rights reserved.
Conflict of interest statement
Conflict of Interest
The authors, Dawn Brasaemle and Carole Stalryd have no conflicts to disclose
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
