

Salt Bridge Formation between the I-BAR Domain and Lipids Increases Lipid Density and Membrane Curvature
- PMID: 28754893
- PMCID: PMC5533756
- DOI: 10.1038/s41598-017-06334-5
Salt Bridge Formation between the I-BAR Domain and Lipids Increases Lipid Density and Membrane Curvature
Abstract
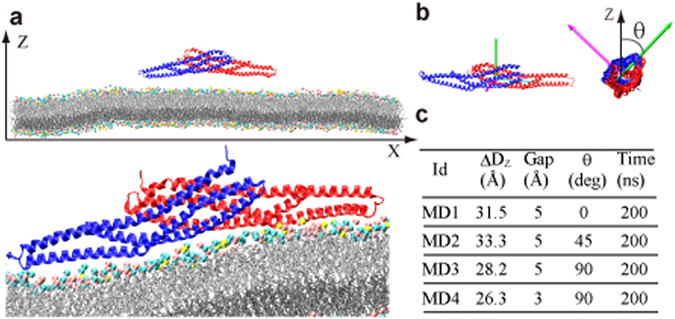
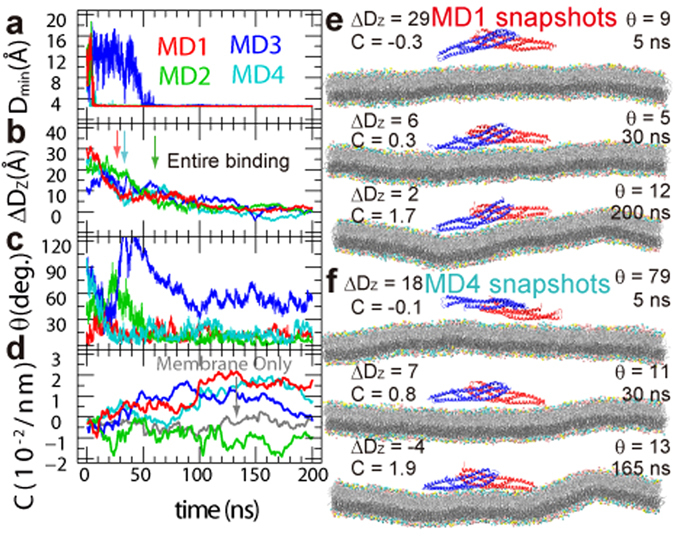
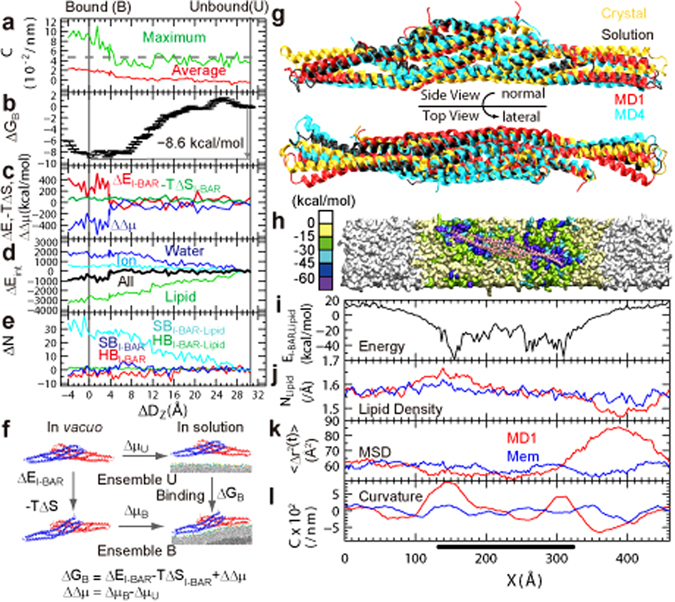
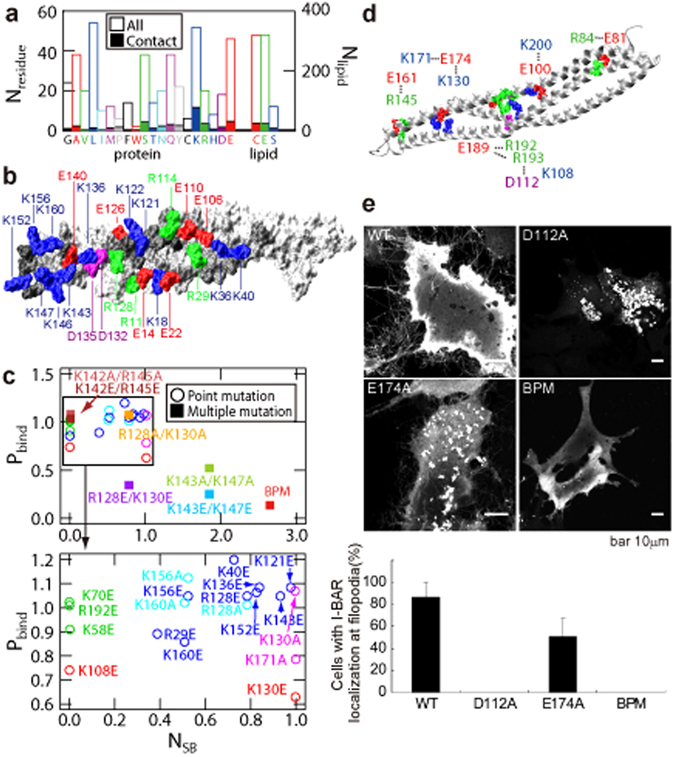
The BAR domain superfamily proteins sense or induce curvature in membranes. The inverse-BAR domain (I-BAR) is a BAR domain that forms a straight "zeppelin-shaped" dimer. The mechanisms by which IRSp53 I-BAR binds to and deforms a lipid membrane are investigated here by all-atom molecular dynamics simulation (MD), binding energy analysis, and the effects of mutation experiments on filopodia on HeLa cells. I-BAR adopts a curved structure when crystallized, but adopts a flatter shape in MD. The binding of I-BAR to membrane was stabilized by ~30 salt bridges, consistent with experiments showing that point mutations of the interface residues have little effect on the binding affinity whereas multiple mutations have considerable effect. Salt bridge formation increases the local density of lipids and deforms the membrane into a concave shape. In addition, the point mutations that break key intra-molecular salt bridges within I-BAR reduce the binding affinity; this was confirmed by expressing these mutants in HeLa cells and observing their effects. The results indicate that the stiffness of I-BAR is important for membrane deformation, although I-BAR does not act as a completely rigid template.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Regulation of membrane-shape transitions induced by I-BAR domains.Biophys J. 2015 Jul 21;109(2):298-307. doi: 10.1016/j.bpj.2015.06.010. Biophys J. 2015. PMID: 26200865 Free PMC article.
-
The RAC binding domain/IRSp53-MIM homology domain of IRSp53 induces RAC-dependent membrane deformation.J Biol Chem. 2006 Nov 17;281(46):35347-58. doi: 10.1074/jbc.M606814200. Epub 2006 Sep 25. J Biol Chem. 2006. PMID: 17003044
-
Molecular mechanisms of membrane deformation by I-BAR domain proteins.Curr Biol. 2009 Jan 27;19(2):95-107. doi: 10.1016/j.cub.2008.12.029. Epub 2009 Jan 15. Curr Biol. 2009. PMID: 19150238
-
Subcellular membrane curvature mediated by the BAR domain superfamily proteins.Semin Cell Dev Biol. 2010 Jun;21(4):340-9. doi: 10.1016/j.semcdb.2009.12.002. Epub 2009 Dec 4. Semin Cell Dev Biol. 2010. PMID: 19963073 Review.
-
I-BAR domains, IRSp53 and filopodium formation.Semin Cell Dev Biol. 2010 Jun;21(4):350-6. doi: 10.1016/j.semcdb.2009.11.008. Epub 2009 Nov 11. Semin Cell Dev Biol. 2010. PMID: 19913105 Review.
Cited by
-
Full assembly of HIV-1 particles requires assistance of the membrane curvature factor IRSp53.Elife. 2021 Jun 11;10:e67321. doi: 10.7554/eLife.67321. Elife. 2021. PMID: 34114563 Free PMC article.
-
Insights into Membrane Curvature Sensing and Membrane Remodeling by Intrinsically Disordered Proteins and Protein Regions.J Membr Biol. 2022 Jun;255(2-3):237-259. doi: 10.1007/s00232-022-00237-x. Epub 2022 Apr 22. J Membr Biol. 2022. PMID: 35451616 Free PMC article. Review.
-
Multiscale Simulations of Biological Membranes: The Challenge To Understand Biological Phenomena in a Living Substance.Chem Rev. 2019 May 8;119(9):5607-5774. doi: 10.1021/acs.chemrev.8b00538. Epub 2019 Mar 12. Chem Rev. 2019. PMID: 30859819 Free PMC article.
-
Mechanism of negative membrane curvature generation by I-BAR domains.Structure. 2021 Dec 2;29(12):1440-1452.e4. doi: 10.1016/j.str.2021.07.010. Epub 2021 Sep 13. Structure. 2021. PMID: 34520736 Free PMC article.
-
Molecular Dynamics Reveal Key Steps in BAR-Related Membrane Remodeling.Pathogens. 2024 Oct 15;13(10):902. doi: 10.3390/pathogens13100902. Pathogens. 2024. PMID: 39452773 Free PMC article.
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
