

Statins are ineffective at reducing neuroinflammation or prolonging survival in scrapie-infected mice
- PMID: 28758631
- PMCID: PMC5721927
- DOI: 10.1099/jgv.0.000876
Statins are ineffective at reducing neuroinflammation or prolonging survival in scrapie-infected mice
Abstract
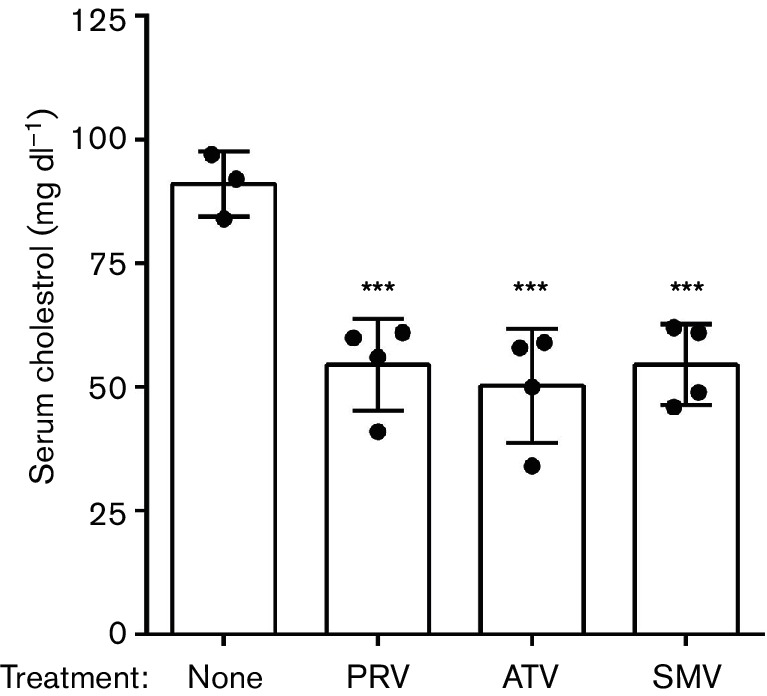
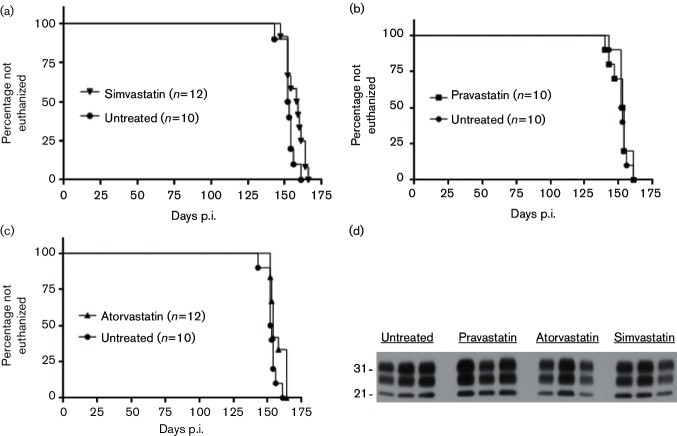
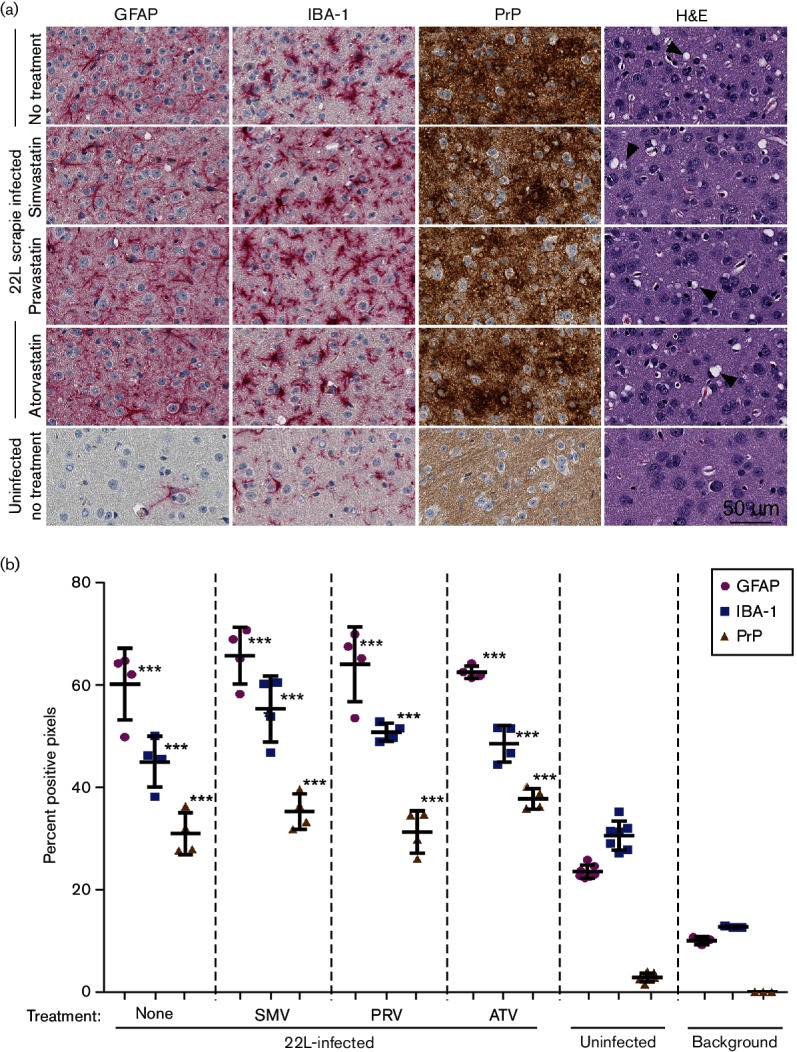
Neuroinflammation is a prominent component of several neurodegenerative diseases, including multiple sclerosis, Alzheimer's disease, Parkinson's disease, tauopathies, amyotrophic lateral sclerosis and prion diseases. In such conditions, the ability to decrease neuroinflammation by drug therapy may influence disease progression. Statins have been used to treat hyperlipidemia as well as reduce neuroinflammation and oxidative stress in various tissues. In previous studies, treatment of scrapie-infected mice with the type 1 statins, simvastatin or pravastatin, showed a small beneficial effect on survival time. In the current study, to increase the effectiveness of statin therapy, we treated infected mice with atorvastatin, a type 2 statin that has improved pharmacokinetics over many type 1 statins. Treatments with either simvastatin or pravastatin were tested for comparison. We evaluated scrapie-infected mice for protease-resistant PrP (PrPres) accumulation, gliosis, neuroinflammation and time until advanced clinical disease requiring euthanasia. All three statin treatments reduced total serum cholesterol ≥40 % in mice. However, gliosis and PrPres deposition were similar in statin-treated and untreated infected mice. Time to euthanasia due to advanced clinical signs was not changed in statin-treated mice relative to untreated mice, a finding at odds with previous reports. Expression of 84 inflammatory genes involved in neuroinflammation was also quantitated. Seven genes were reduced by pravastatin, and one gene was reduced by atorvastatin. In contrast, simvastatin therapy did not reduce any of the tested genes, but did slightly increase the expression of Ccl2 and Cxcl13. Our studies indicate that none of the three statins tested were effective in reducing scrapie-induced neuroinflammation or neuropathogenesis.
Figures
Similar articles
-
Prion infection of mouse brain reveals multiple new upregulated genes involved in neuroinflammation or signal transduction.J Virol. 2015 Feb;89(4):2388-404. doi: 10.1128/JVI.02952-14. Epub 2014 Dec 10. J Virol. 2015. PMID: 25505076 Free PMC article.
-
Simvastatin treatment prolongs the survival of scrapie-infected mice.Neuroreport. 2007 Mar 26;18(5):479-82. doi: 10.1097/WNR.0b013e328058678d. Neuroreport. 2007. PMID: 17496807
-
Redox markers and inflammation are differentially affected by atorvastatin, pravastatin or simvastatin administered before endotoxin-induced acute lung injury.Int Immunopharmacol. 2013 Sep;17(1):57-64. doi: 10.1016/j.intimp.2013.05.016. Epub 2013 Jun 6. Int Immunopharmacol. 2013. PMID: 23747588
-
Are statins created equal? Evidence from randomized trials of pravastatin, simvastatin, and atorvastatin for cardiovascular disease prevention.Am Heart J. 2006 Feb;151(2):273-81. doi: 10.1016/j.ahj.2005.04.003. Am Heart J. 2006. PMID: 16442888 Review.
-
Atorvastatin: gold standard for prophylaxis of myocardial ischemia and stroke - comparison of the clinical benefit of statins on the basis of randomized controlled endpoint studies.Eur J Med Res. 2004 Jan 26;9(1):1-17. Eur J Med Res. 2004. PMID: 14766335 Review.
Cited by
-
Transient receptor potential vanilloid channel 2 contributes to multi-modal endoplasmic reticulum and perinuclear space dilations that can also be observed in prion-infected mice.Acta Neuropathol. 2025 Jun 23;149(1):68. doi: 10.1007/s00401-025-02906-2. Acta Neuropathol. 2025. PMID: 40549026
-
A unified model for the origins of spongiform degeneration and other neuropathological features in prion diseases.ArXiv [Preprint]. 2025 Jan 15:arXiv:2412.16678v2. ArXiv. 2025. PMID: 39876936 Free PMC article. Preprint.
-
The Antineuroinflammatory Effect of Simvastatin on Lipopolysaccharide Activated Microglial Cells.Evid Based Complement Alternat Med. 2018 Nov 7;2018:9691085. doi: 10.1155/2018/9691085. eCollection 2018. Evid Based Complement Alternat Med. 2018. PMID: 30524484 Free PMC article.
-
Absence of Apolipoprotein E is associated with exacerbation of prion pathology and promotes microglial neurodegenerative phenotype.Acta Neuropathol Commun. 2021 Sep 26;9(1):157. doi: 10.1186/s40478-021-01261-z. Acta Neuropathol Commun. 2021. PMID: 34565486 Free PMC article.
-
Neuroinflammation, Microglia, and Cell-Association during Prion Disease.Viruses. 2019 Jan 15;11(1):65. doi: 10.3390/v11010065. Viruses. 2019. PMID: 30650564 Free PMC article. Review.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials