

Variation in Extracellular Detoxification Is a Link to Different Carcinogenicity among Chromates in Rodent and Human Lungs
- PMID: 28759204
- PMCID: PMC5605882
- DOI: 10.1021/acs.chemrestox.7b00172
Variation in Extracellular Detoxification Is a Link to Different Carcinogenicity among Chromates in Rodent and Human Lungs
Abstract
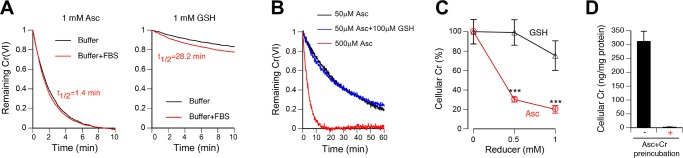
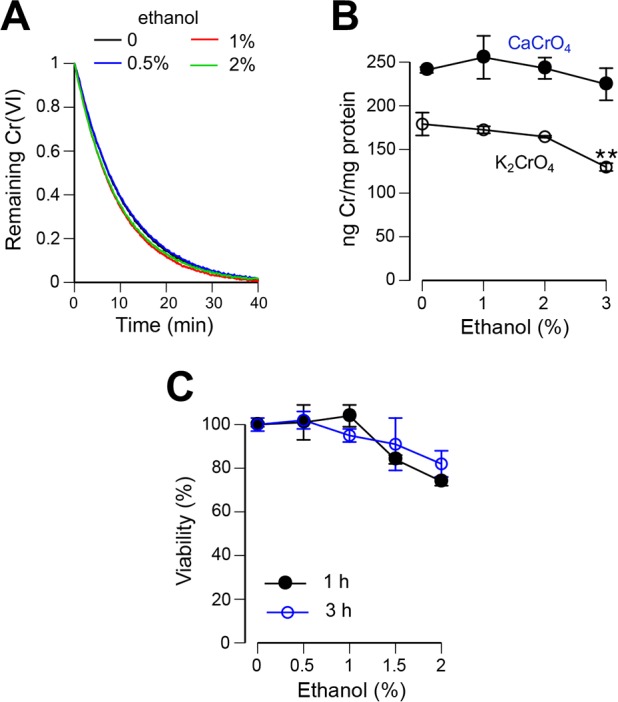
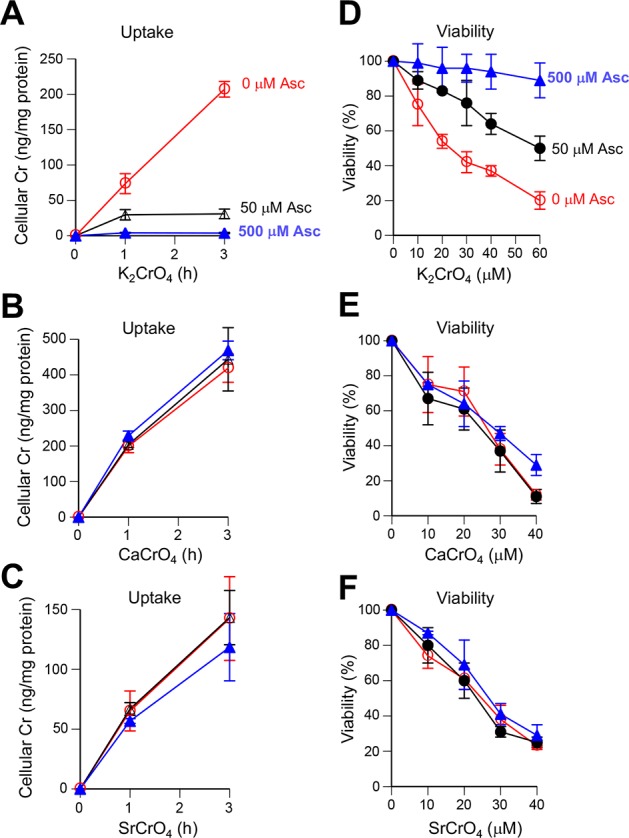
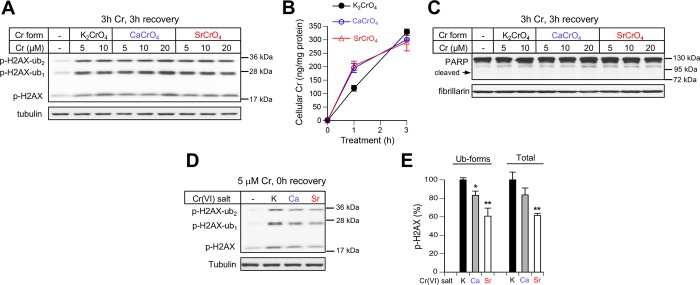
Inhalation of soluble chromium(VI) is firmly linked with higher risks of lung cancer in humans. However, comparative studies in rats have found a high lung tumorigenicity for moderately soluble chromates but no tumors for highly soluble chromates. These major species differences remain unexplained. We investigated the impact of extracellular reducers on responses of human and rat lung epithelial cells to different Cr(VI) forms. Extracellular reduction of Cr(VI) is a detoxification process, and rat and human lung lining fluids contain different concentrations of ascorbate and glutathione. We found that reduction of chromate anions in simulated lung fluids was principally driven by ascorbate with only minimal contribution from glutathione. The addition of 500 μM ascorbate (∼rat lung fluid concentration) to culture media strongly inhibited cellular uptake of chromate anions and completely prevented their cytotoxicity even at otherwise lethal doses. While proportionally less effective, 50 μM extracellular ascorbate (∼human lung fluid concentration) also decreased uptake of chromate anions and their cytotoxicity. In comparison to chromate anions, uptake and cytotoxicity of respirable particles of moderately soluble CaCrO4 and SrCrO4 were much less sensitive to suppression by extracellular ascorbate, especially during early exposure times and in primary bronchial cells. In the absence of extracellular ascorbate, chromate anions and CaCrO4/SrCrO4 particles produced overall similar levels of DNA double-stranded breaks, with less soluble particles exhibiting a slower rate of breakage. Our results indicate that a gradual extracellular dissolution and a rapid internalization of calcium chromate and strontium chromate particles makes them resistant to detoxification outside the cells, which is extremely effective for chromate anions in the rat lung fluid. The detoxification potential of the human lung fluid is significant but much lower and insufficient to provide a threshold-type dose dependence for soluble chromates.
Conflict of interest statement
The authors declare no competing financial interest.
Figures





Similar articles
-
Toxicological Antagonism among Welding Fume Metals: Inactivation of Soluble Cr(VI) by Iron.Chem Res Toxicol. 2018 Nov 19;31(11):1172-1184. doi: 10.1021/acs.chemrestox.8b00182. Epub 2018 Nov 6. Chem Res Toxicol. 2018. PMID: 30362728 Free PMC article.
-
Lead chromate-induced chromosome damage requires extracellular dissolution to liberate chromium ions but does not require particle internalization or intracellular dissolution.Chem Res Toxicol. 2004 Oct;17(10):1362-7. doi: 10.1021/tx0498509. Chem Res Toxicol. 2004. PMID: 15487897
-
Lead ions do not cause human lung cells to escape chromate-induced cytotoxicity.Toxicol Appl Pharmacol. 2005 Mar 1;203(2):167-76. doi: 10.1016/j.taap.2004.08.006. Toxicol Appl Pharmacol. 2005. PMID: 15710177
-
The role of chromium(V) in the mechanism of chromate-induced oxidative DNA damage and cancer.J Environ Pathol Toxicol Oncol. 2000;19(3):215-30. J Environ Pathol Toxicol Oncol. 2000. PMID: 10983888 Review.
-
Plasmid chromate resistance and chromate reduction.Plasmid. 1992 Jan;27(1):65-71. doi: 10.1016/0147-619x(92)90007-w. Plasmid. 1992. PMID: 1741461 Review.
Cited by
-
Chemical mechanisms of DNA damage by carcinogenic chromium(VI).Adv Pharmacol. 2023;96:25-46. doi: 10.1016/bs.apha.2022.07.003. Epub 2022 Aug 26. Adv Pharmacol. 2023. PMID: 36858775 Free PMC article.
-
Nuclear SUMOylation and Proteotoxic Stress Responses to Metals with Different Ligand Preferences.Chem Res Toxicol. 2025 May 19;38(5):942-953. doi: 10.1021/acs.chemrestox.5c00040. Epub 2025 Apr 17. Chem Res Toxicol. 2025. PMID: 40243484
-
Toxicological Antagonism among Welding Fume Metals: Inactivation of Soluble Cr(VI) by Iron.Chem Res Toxicol. 2018 Nov 19;31(11):1172-1184. doi: 10.1021/acs.chemrestox.8b00182. Epub 2018 Nov 6. Chem Res Toxicol. 2018. PMID: 30362728 Free PMC article.
-
Effects of Sodium Chromate Exposure on Gene Expression Profiles of Primary Rat Hepatocytes (In Vitro).Biol Trace Elem Res. 2023 Apr;201(4):1913-1934. doi: 10.1007/s12011-022-03294-4. Epub 2022 Jun 2. Biol Trace Elem Res. 2023. PMID: 35653032
-
Nuclear and Cytoplasmic Functions of Vitamin C.Chem Res Toxicol. 2020 Oct 19;33(10):2515-2526. doi: 10.1021/acs.chemrestox.0c00348. Epub 2020 Oct 1. Chem Res Toxicol. 2020. PMID: 33001635 Free PMC article. Review.
References
-
- Costa M.; Zhitkovich A.; Gargas M.; Paustenbach D.; Finley B.; Kuykendall J.; Billings R.; Carlson T.; Wetterhahn K.; Xu J.; Patierno S.; Bogdanffy M. (1996) Interlaboratory validation of a new assay for DNA-protein crosslinks. Mutat. Res., Genet. Toxicol. Test. 369, 13–21. 10.1016/S0165-1218(96)90043-9. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
