

Rethinking m6A Readers, Writers, and Erasers
- PMID: 28759256
- PMCID: PMC5963928
- DOI: 10.1146/annurev-cellbio-100616-060758
Rethinking m6A Readers, Writers, and Erasers
Abstract
In recent years, m6A has emerged as an abundant and dynamically regulated modification throughout the transcriptome. Recent technological advances have enabled the transcriptome-wide identification of m6A residues, which in turn has provided important insights into the biology and regulation of this pervasive regulatory mark. Also central to our current understanding of m6A are the discovery and characterization of m6A readers, writers, and erasers. Over the last few years, studies into the function of these proteins have led to important discoveries about the regulation and function of m6A. However, during this time our understanding of these proteins has also evolved considerably, sometimes leading to the reversal of early concepts regarding the reading, writing and erasing of m6A. In this review, we summarize recent advances in m6A research, and we highlight how these new findings have reshaped our understanding of how m6A is regulated in the transcriptome.
Keywords: RNA modifications; epitranscriptome; m6A; posttranscriptional control.
Figures
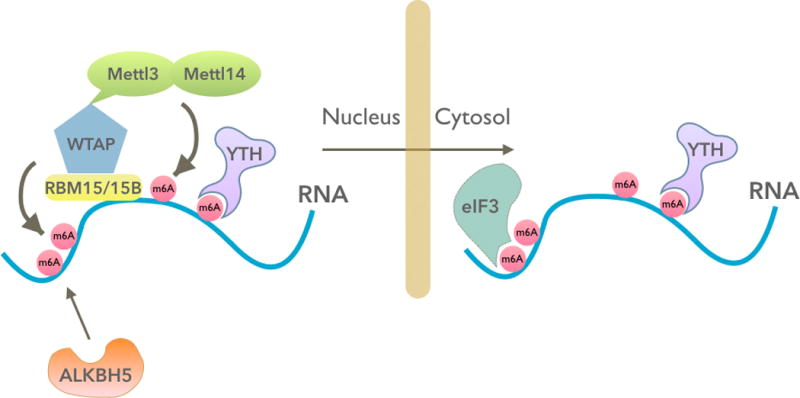
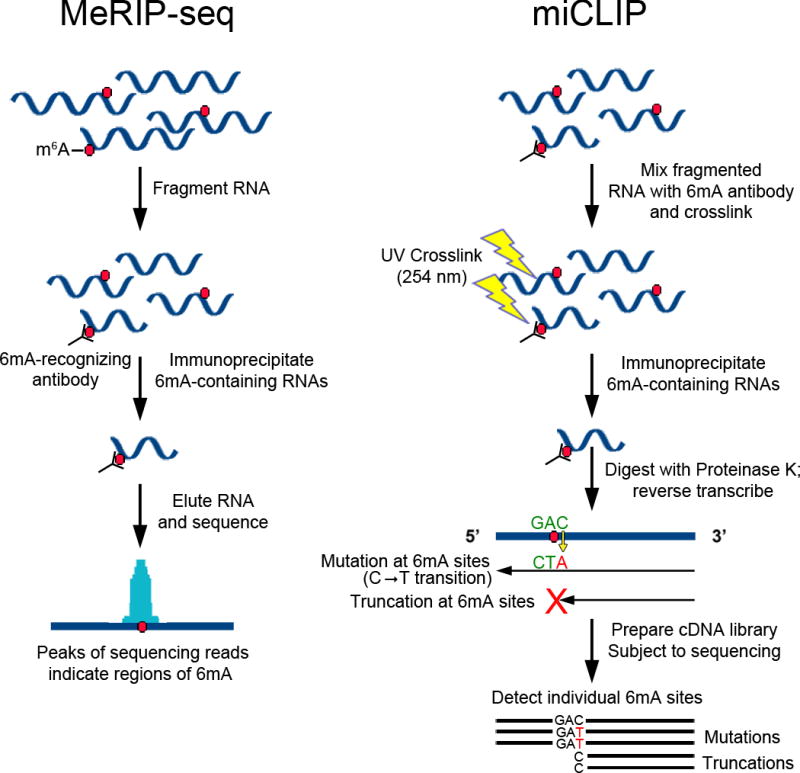
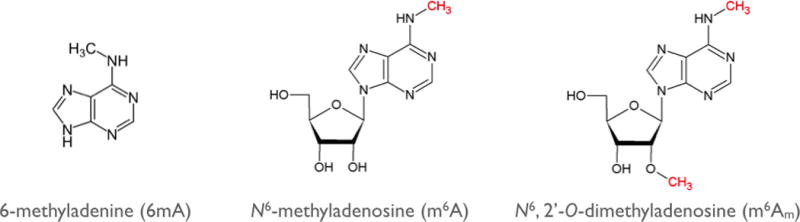
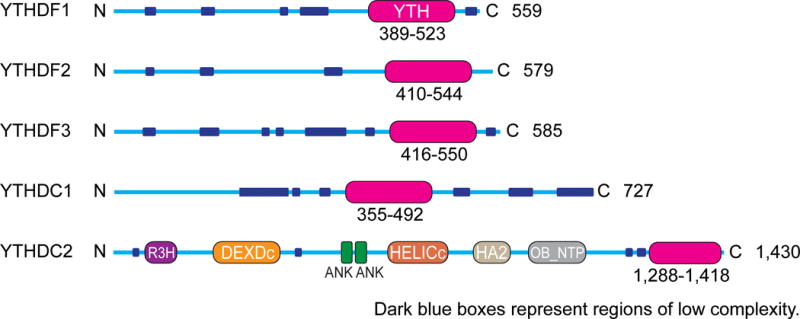
References
-
- Aguzzi A, Altmeyer M. Phase separation: linking cellular compartmentalization to disease. Trends Cell Biol. 2016;26:547–58. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
