

An acetylation-phosphorylation switch that regulates tau aggregation propensity and function
- PMID: 28760828
- PMCID: PMC5602388
- DOI: 10.1074/jbc.M117.794602
An acetylation-phosphorylation switch that regulates tau aggregation propensity and function
Abstract
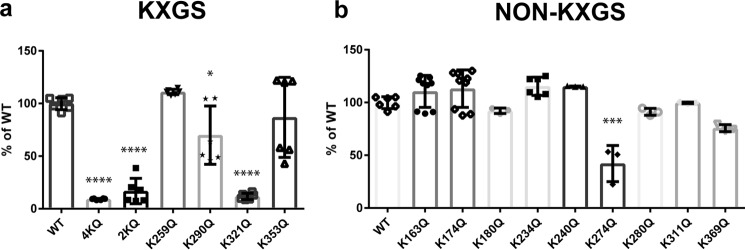
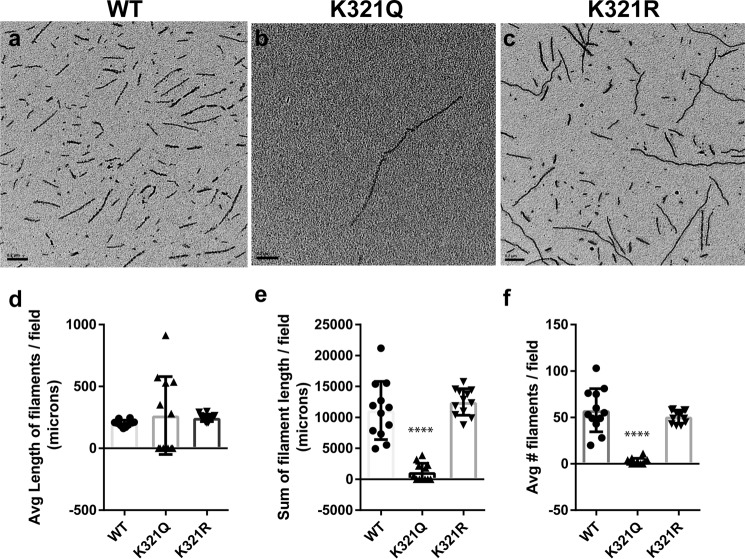
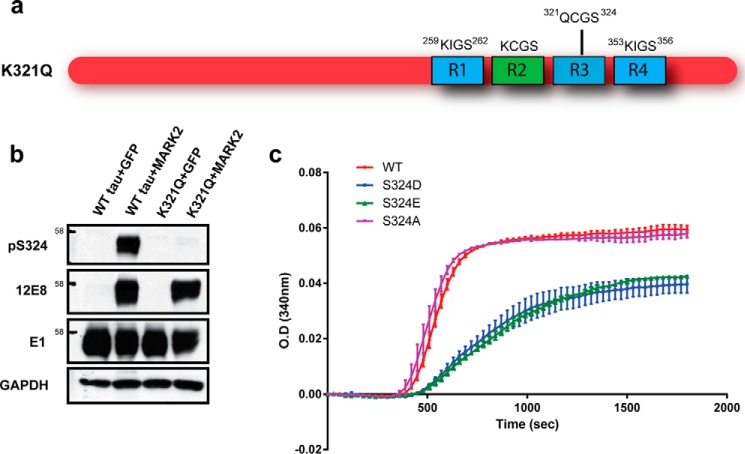
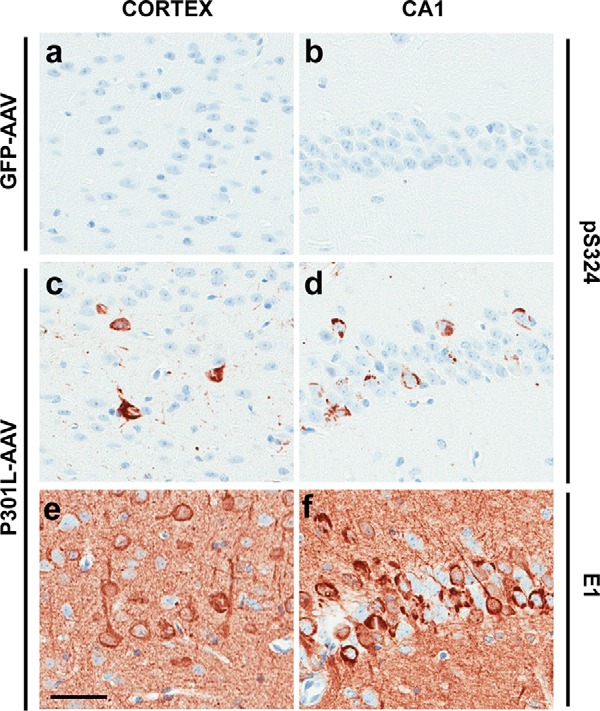
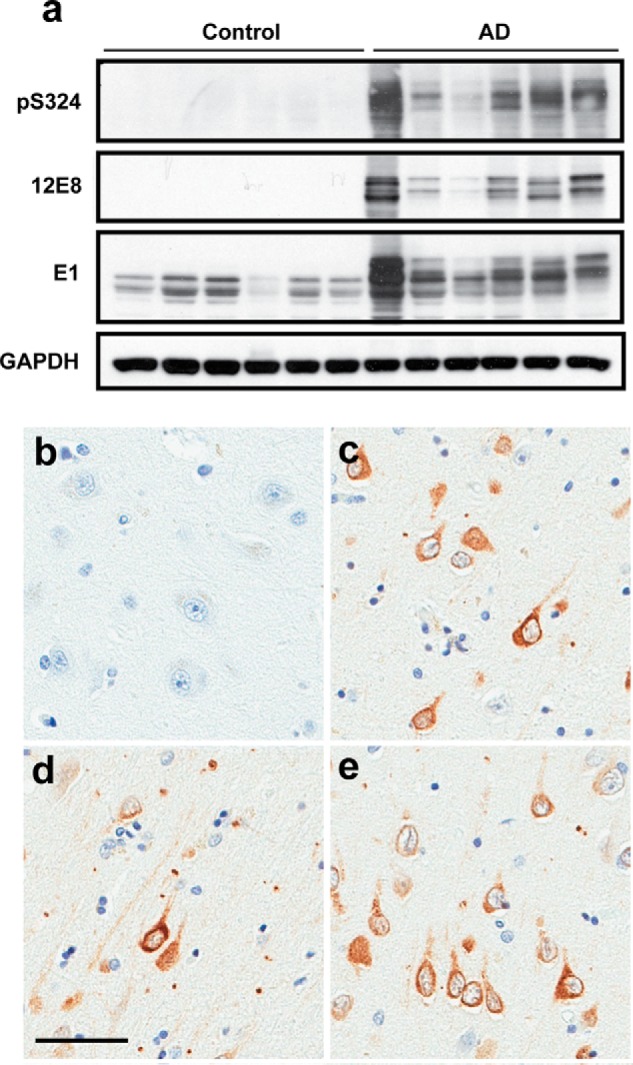
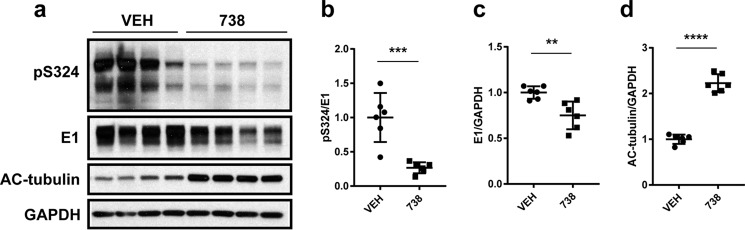
The aberrant accumulation of tau protein is a pathological hallmark of a class of neurodegenerative diseases known as tauopathies, including Alzheimer's disease and related dementias. On the basis of previous observations that tau is a direct substrate of histone deacetylase 6 (HDAC6), we sought to map all HDAC6-responsive sites in tau and determine how acetylation in a site-specific manner affects tau's biophysical properties in vitro Our findings indicate that several acetylation sites in tau are responsive to HDAC6 and that acetylation on Lys-321 (within a KCGS motif) is both essential for acetylation-mediated inhibition of tau aggregation in vitro and a molecular tactic for preventing phosphorylation on the downstream Ser-324 residue. To determine the functional consequence of this HDAC6-regulated phosphorylation event, we examined tau's ability to promote microtubule assembly and found that phosphorylation of Ser-324 interferes with the normal microtubule-stabilizing function of tau. Tau phosphorylation of Ser-324 (pSer-324) has not previously been evaluated in the context of tauopathy, and here we observed increased deposition of pSer-324-positive tau both in mouse models of tauopathy and in patients with Alzheimer's disease. These findings uncover a novel acetylation-phosphorylation switch at Lys-321/Ser-324 that coordinately regulates tau polymerization and function. Because the disease relevance of this finding is evident, additional studies are needed to examine the role of pSer-324 in tau pathobiology and to determine whether therapeutically modulating this acetylation-phosphorylation switch affects disease progression in vivo.
Keywords: Alzheimer disease; Tau protein (Tau); acetylation; aggregation; histone deacetylase 6 (HDAC6); neurodegenerative disease; phosphorylation; tauopathy.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Buée L., Bussière T., Buée-Scherrer V., Delacourte A., and Hof P. R. (2000) Tau protein isoforms, phosphorylation and role in neurodegenerative disorders. Brain Res. Brain Res. Rev. 33, 95–130 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
