

RORγt and RORα signature genes in human Th17 cells
- PMID: 28763457
- PMCID: PMC5538713
- DOI: 10.1371/journal.pone.0181868
RORγt and RORα signature genes in human Th17 cells
Abstract
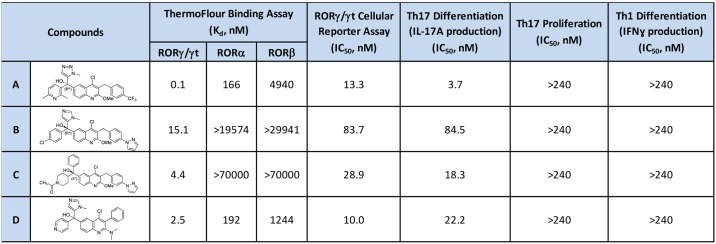
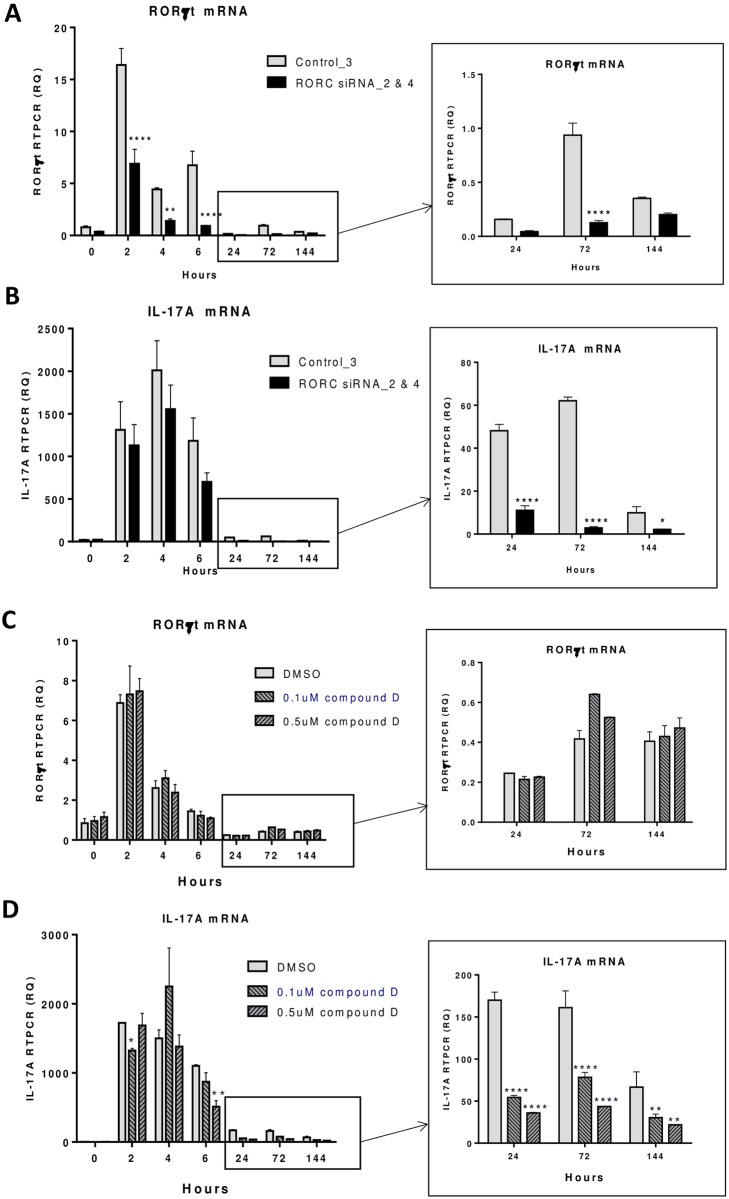
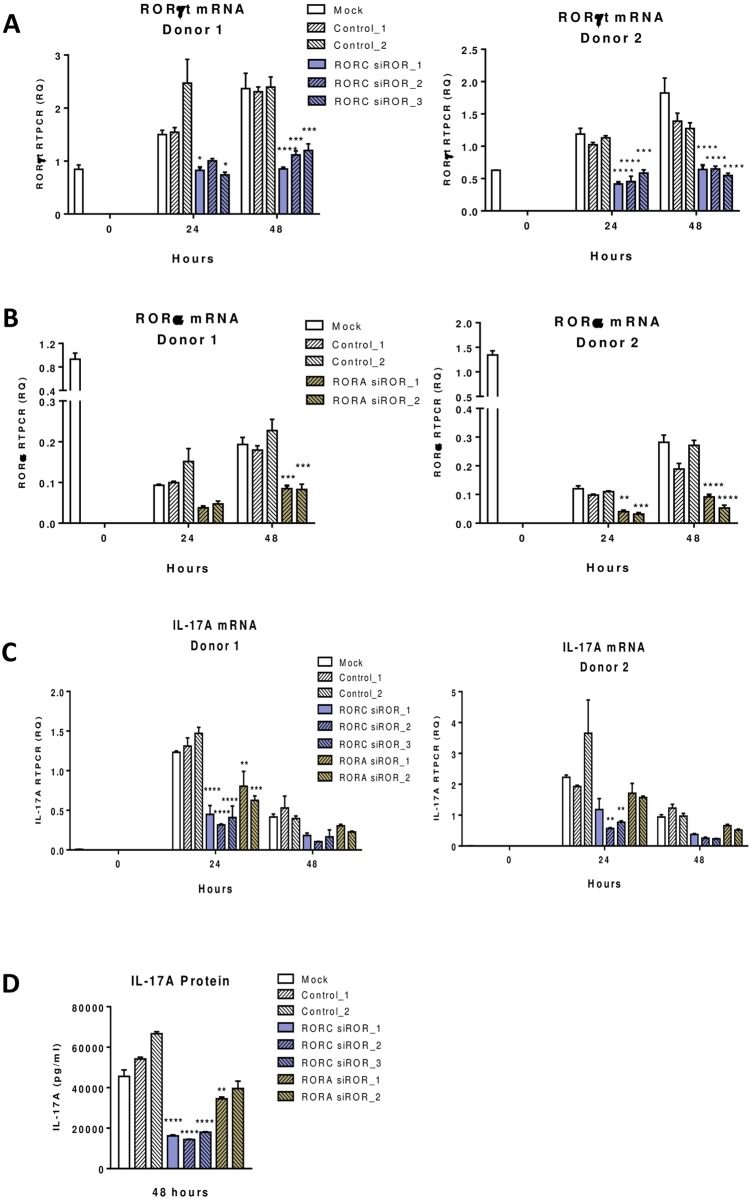
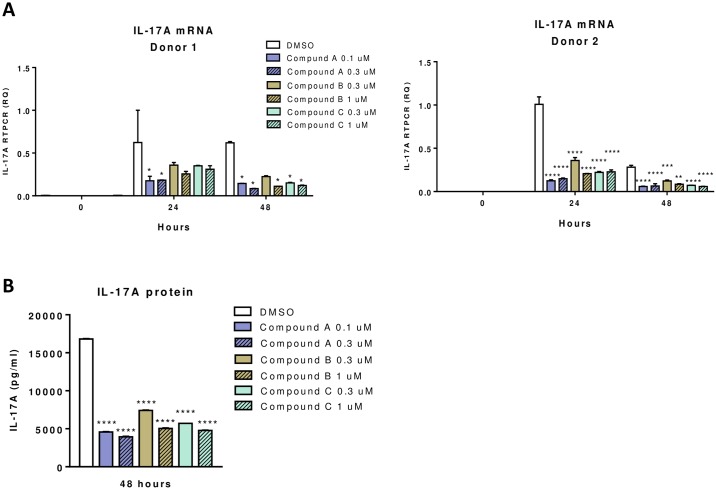
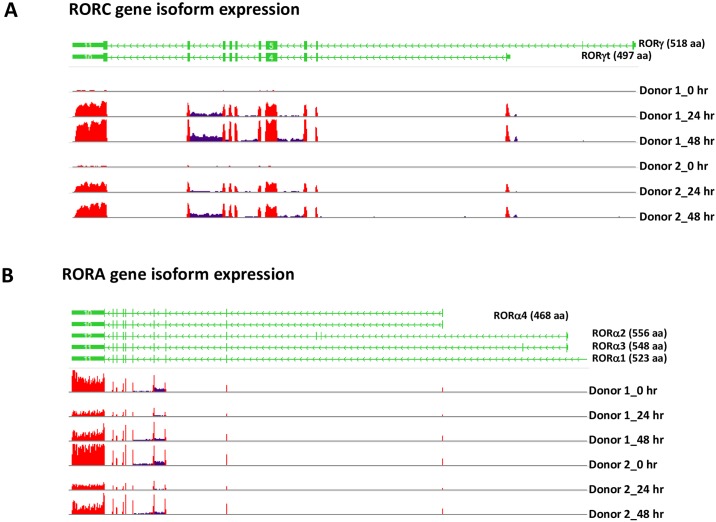
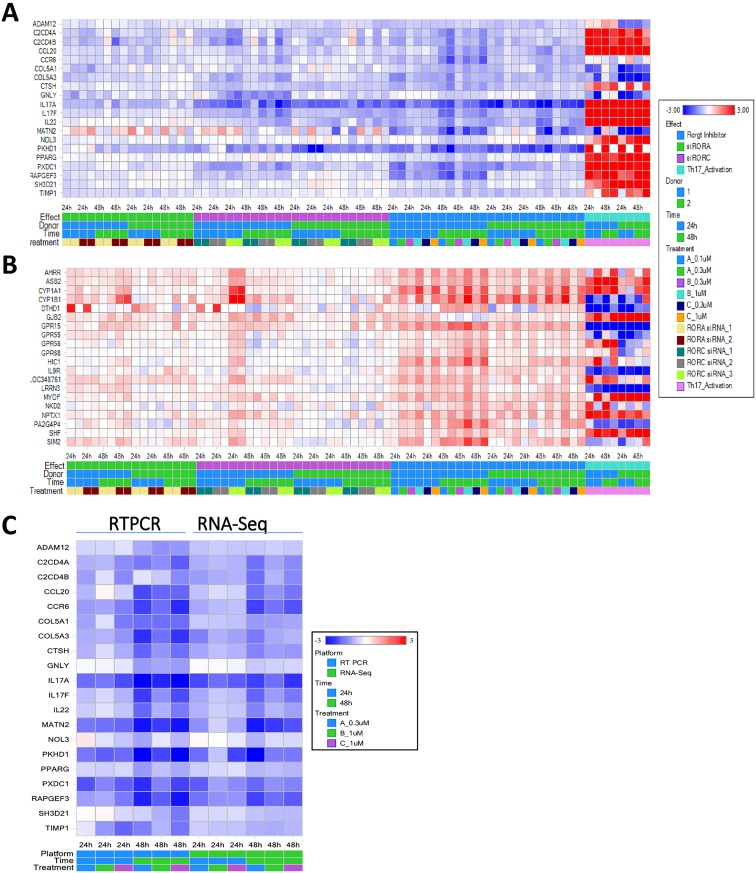
RORγt and RORα are transcription factors of the RAR-related orphan nuclear receptor (ROR) family. They are expressed in Th17 cells and have been suggested to play a role in Th17 differentiation. Although RORγt signature genes have been characterized in mouse Th17 cells, detailed information on its transcriptional control in human Th17 cells is limited and even less is known about RORα signature genes which have not been reported in either human or mouse T cells. In this study, global gene expression of human CD4 T cells activated under Th17 skewing conditions was profiled by RNA sequencing. RORγt and RORα signature genes were identified in these Th17 cells treated with specific siRNAs to knock down RORγt or RORα expression. We have generated selective small molecule RORγt modulators and they were also utilized as pharmacological tools in RORγt signature gene identification. Our results showed that RORγt controlled the expression of a very selective number of genes in Th17 cells and most of them were regulated by RORα as well albeit a weaker influence. Key Th17 genes including IL-17A, IL-17F, IL-23R, CCL20 and CCR6 were shown to be regulated by both RORγt and RORα. Our results demonstrated an overlapping role of RORγt and RORα in human Th17 cell differentiation through regulation of a defined common set of Th17 genes. RORγt as a drug target for treatment of Th17 mediated autoimmune diseases such as psoriasis has been demonstrated recently in clinical trials. Our results suggest that RORα could be involved in same disease mechanisms and gene signatures identified in this report could be valuable biomarkers for tracking the pharmacodynamic effects of compounds that modulate RORγt or RORα activities in patients.
Conflict of interest statement
Figures

References
-
- Jetten AM (2009) Retinoid-related orphan receptors (RORs): critical roles in development, immunity, circadian rhythm, and cellular metabolism. Nucl Recept Signal 7: e003 doi: 10.1621/nrs.07003 - DOI - PMC - PubMed
-
- Giguere V, Tini M, Flock G, Ong E, Evans RM, et al. (1994) Isoform-specific amino-terminal domains dictate DNA-binding properties of ROR alpha, a novel family of orphan hormone nuclear receptors. Genes Dev 8: 538–553. - PubMed
-
- Matysiak-Scholze U, Nehls M (1997) The structural integrity of ROR alpha isoforms is mutated in staggerer mice: cerebellar coexpression of ROR alpha1 and ROR alpha4. Genomics 43: 78–84. - PubMed
-
- Carlberg C, Hooft van Huijsduijnen R, Staple JK, DeLamarter JF, Becker-Andre M (1994) RZRs, a new family of retinoid-related orphan receptors that function as both monomers and homodimers. Mol Endocrinol 8: 757–770. - PubMed
-
- Villey I, de Chasseval R, de Villartay JP (1999) RORgammaT, a thymus-specific isoform of the orphan nuclear receptor RORgamma / TOR, is up-regulated by signaling through the pre-T cell receptor and binds to the TEA promoter. Eur J Immunol 29: 4072–4080. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
