

Methane-Fueled Syntrophy through Extracellular Electron Transfer: Uncovering the Genomic Traits Conserved within Diverse Bacterial Partners of Anaerobic Methanotrophic Archaea
- PMID: 28765215
- PMCID: PMC5539420
- DOI: 10.1128/mBio.00530-17
Methane-Fueled Syntrophy through Extracellular Electron Transfer: Uncovering the Genomic Traits Conserved within Diverse Bacterial Partners of Anaerobic Methanotrophic Archaea
Erratum in
-
Erratum for Skennerton et al., "Methane-Fueled Syntrophy through Extracellular Electron Transfer: Uncovering the Genomic Traits Conserved within Diverse Bacterial Partners of Anaerobic Methanotrophic Archaea".mBio. 2017 Oct 3;8(5):e01561-17. doi: 10.1128/mBio.01561-17. mBio. 2017. PMID: 28974619 Free PMC article. No abstract available.
Abstract
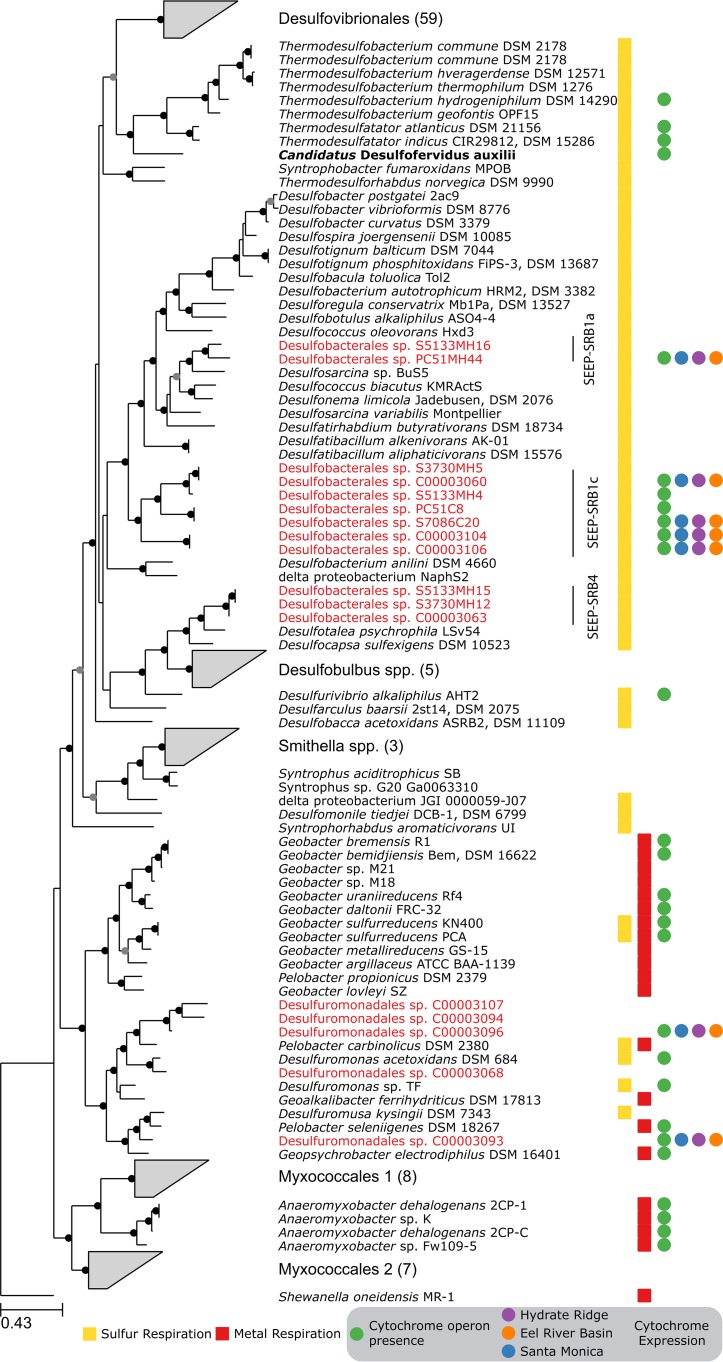
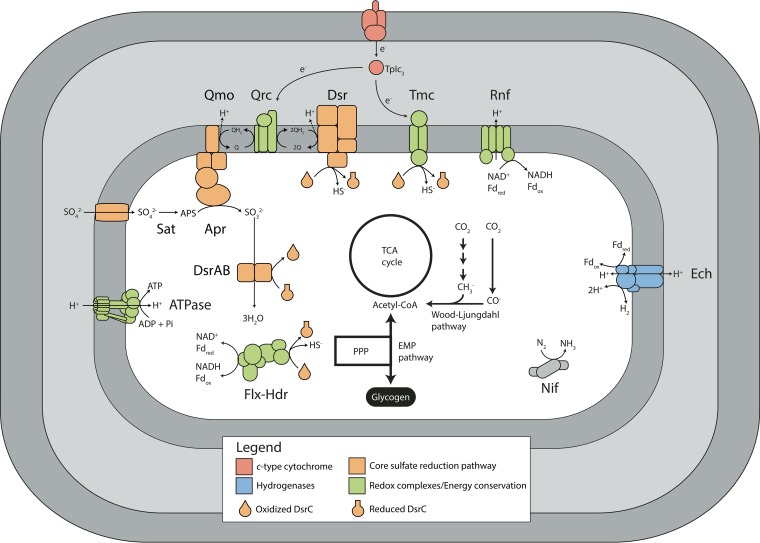
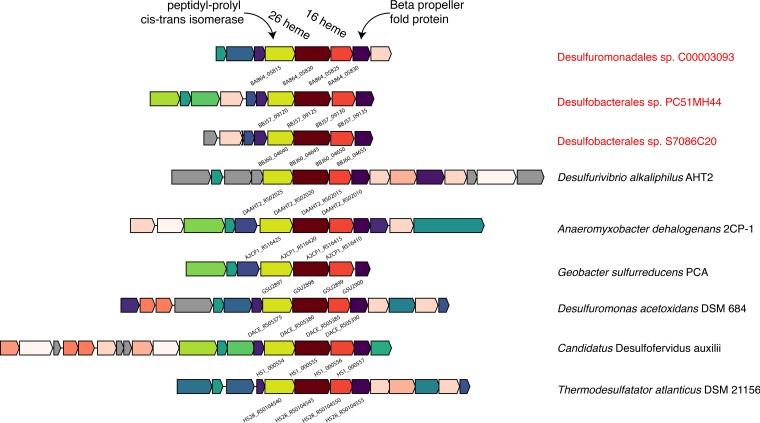
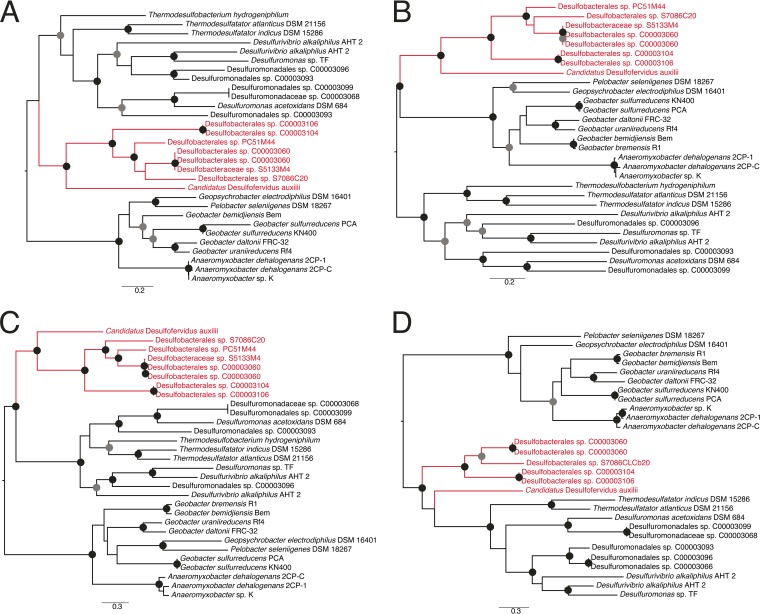
The anaerobic oxidation of methane by anaerobic methanotrophic (ANME) archaea in syntrophic partnership with deltaproteobacterial sulfate-reducing bacteria (SRB) is the primary mechanism for methane removal in ocean sediments. The mechanism of their syntrophy has been the subject of much research as traditional intermediate compounds, such as hydrogen and formate, failed to decouple the partners. Recent findings have indicated the potential for extracellular electron transfer from ANME archaea to SRB, though it is unclear how extracellular electrons are integrated into the metabolism of the SRB partner. We used metagenomics to reconstruct eight genomes from the globally distributed SEEP-SRB1 clade of ANME partner bacteria to determine what genomic features are required for syntrophy. The SEEP-SRB1 genomes contain large multiheme cytochromes that were not found in previously described free-living SRB and also lack periplasmic hydrogenases that may prevent an independent lifestyle without an extracellular source of electrons from ANME archaea. Metaproteomics revealed the expression of these cytochromes at in situ methane seep sediments from three sites along the Pacific coast of the United States. Phylogenetic analysis showed that these cytochromes appear to have been horizontally transferred from metal-respiring members of the Deltaproteobacteria such as Geobacter and may allow these syntrophic SRB to accept extracellular electrons in place of other chemical/organic electron donors.IMPORTANCE Some archaea, known as anaerobic methanotrophs, are capable of converting methane into carbon dioxide when they are growing syntopically with sulfate-reducing bacteria. This partnership is the primary mechanism for methane removal in ocean sediments; however, there is still much to learn about how this syntrophy works. Previous studies have failed to identify the metabolic intermediate, such as hydrogen or formate, that is passed between partners. However, recent analysis of methanotrophic archaea has suggested that the syntrophy is formed through direct electron transfer. In this research, we analyzed the genomes of multiple partner bacteria and showed that they also contain the genes necessary to perform extracellular electron transfer, which are absent in related bacteria that do not form syntrophic partnerships with anaerobic methanotrophs. This genomic evidence shows a possible mechanism for direct electron transfer from methanotrophic archaea into the metabolism of the partner bacteria.
Keywords: ANME; AOM; SEEP-SRB1; anaerobic oxidation of methane; extracellular electron transfer; methane seeps; multiheme cytochrome; sulfate-reducing bacteria.
Copyright © 2017 Skennerton et al.
Figures
Similar articles
-
Comparative genomics reveals electron transfer and syntrophic mechanisms differentiating methanotrophic and methanogenic archaea.PLoS Biol. 2022 Jan 5;20(1):e3001508. doi: 10.1371/journal.pbio.3001508. eCollection 2022 Jan. PLoS Biol. 2022. PMID: 34986141 Free PMC article. Review.
-
Community Structure and Microbial Associations in Sediment-Free Methanotrophic Enrichment Cultures from a Marine Methane Seep.Appl Environ Microbiol. 2022 Jun 14;88(11):e0210921. doi: 10.1128/aem.02109-21. Epub 2022 May 23. Appl Environ Microbiol. 2022. PMID: 35604226 Free PMC article.
-
Intercellular wiring enables electron transfer between methanotrophic archaea and bacteria.Nature. 2015 Oct 22;526(7574):587-90. doi: 10.1038/nature15733. Nature. 2015. PMID: 26490622
-
Physiological potential and evolutionary trajectories of syntrophic sulfate-reducing bacterial partners of anaerobic methanotrophic archaea.PLoS Biol. 2023 Sep 25;21(9):e3002292. doi: 10.1371/journal.pbio.3002292. eCollection 2023 Sep. PLoS Biol. 2023. PMID: 37747940 Free PMC article.
-
Genomic Insights into Niche Partitioning across Sediment Depth among Anaerobic Methane-Oxidizing Archaea in Global Methane Seeps.mSystems. 2023 Apr 27;8(2):e0117922. doi: 10.1128/msystems.01179-22. Epub 2023 Mar 16. mSystems. 2023. PMID: 36927099 Free PMC article. Review.
Cited by
-
Comparative genomics reveals electron transfer and syntrophic mechanisms differentiating methanotrophic and methanogenic archaea.PLoS Biol. 2022 Jan 5;20(1):e3001508. doi: 10.1371/journal.pbio.3001508. eCollection 2022 Jan. PLoS Biol. 2022. PMID: 34986141 Free PMC article. Review.
-
The Proposed Molecular Mechanisms Used by Archaea for Fe(III) Reduction and Fe(II) Oxidation.Front Microbiol. 2021 Jul 1;12:690918. doi: 10.3389/fmicb.2021.690918. eCollection 2021. Front Microbiol. 2021. PMID: 34276623 Free PMC article. Review.
-
Chemoorganoautotrophic lifestyle of the anaerobic enrichment culture N47 growing on naphthalene.Commun Biol. 2025 Jun 4;8(1):856. doi: 10.1038/s42003-025-08172-y. Commun Biol. 2025. PMID: 40467871 Free PMC article.
-
Lateral Gene Transfer Drives Metabolic Flexibility in the Anaerobic Methane-Oxidizing Archaeal Family Methanoperedenaceae.mBio. 2020 Jun 30;11(3):e01325-20. doi: 10.1128/mBio.01325-20. mBio. 2020. PMID: 32605988 Free PMC article.
-
Identification of a Redox Active Thioquinoxalinol Sulfate Compound Produced by an Anaerobic Methane-Oxidizing Microbial Consortium.ACS Omega. 2019 Dec 16;4(27):22613-22622. doi: 10.1021/acsomega.9b03450. eCollection 2019 Dec 31. ACS Omega. 2019. PMID: 31909345 Free PMC article.
References
-
- Boetius A, Wenzhöfer F. 2013. Seafloor oxygen consumption fuelled by methane from cold seeps. Nat Geosci 6:725–734. doi:10.1038/ngeo1926. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
