

Calcium Ion Induced Structural Changes Promote Dimerization of Secretagogin, Which Is Required for Its Insulin Secretory Function
- PMID: 28765527
- PMCID: PMC5539292
- DOI: 10.1038/s41598-017-07072-4
Calcium Ion Induced Structural Changes Promote Dimerization of Secretagogin, Which Is Required for Its Insulin Secretory Function
Abstract
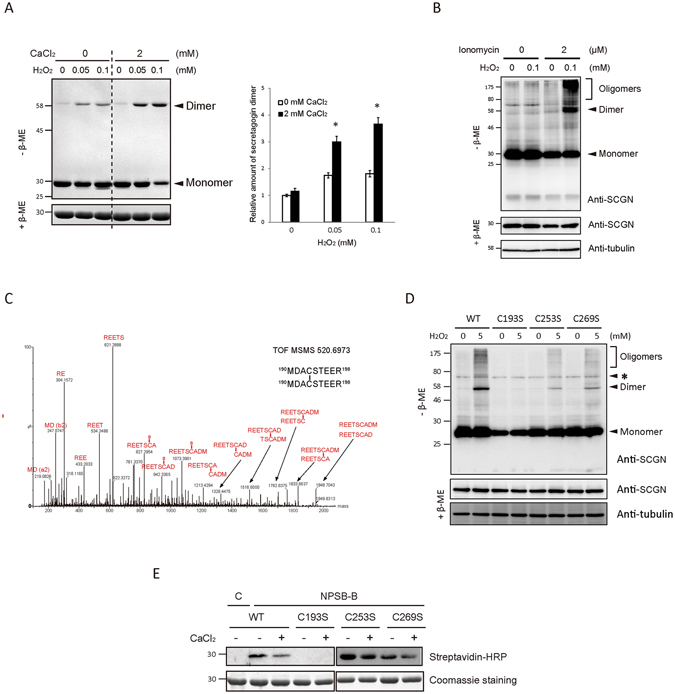
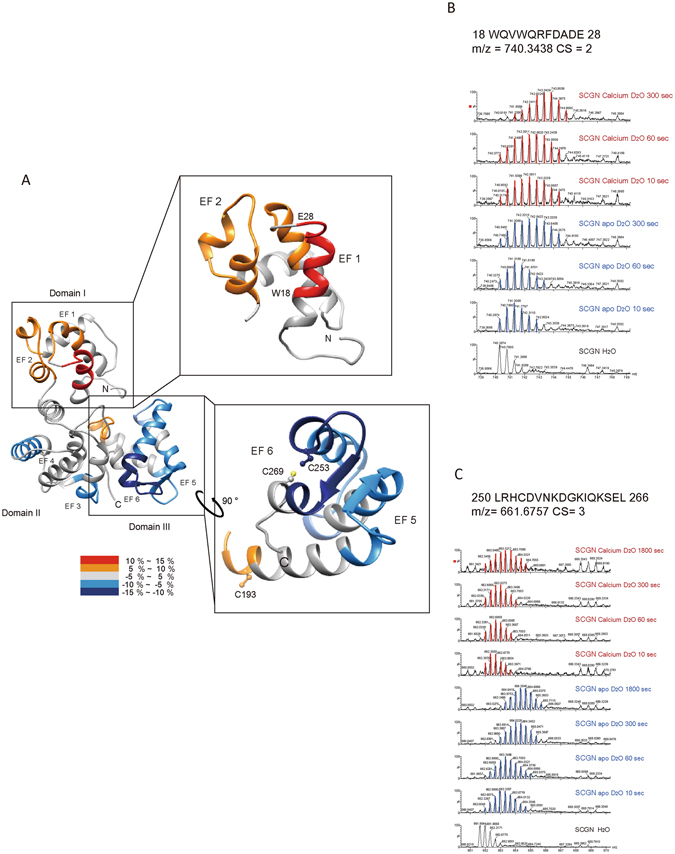
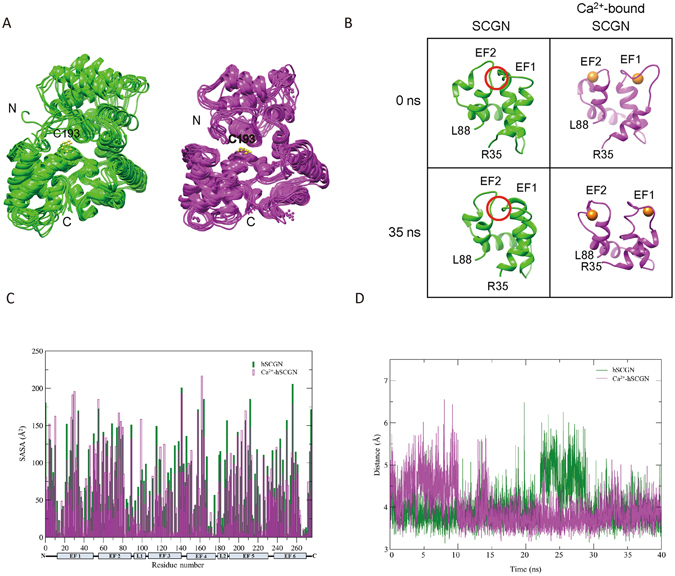
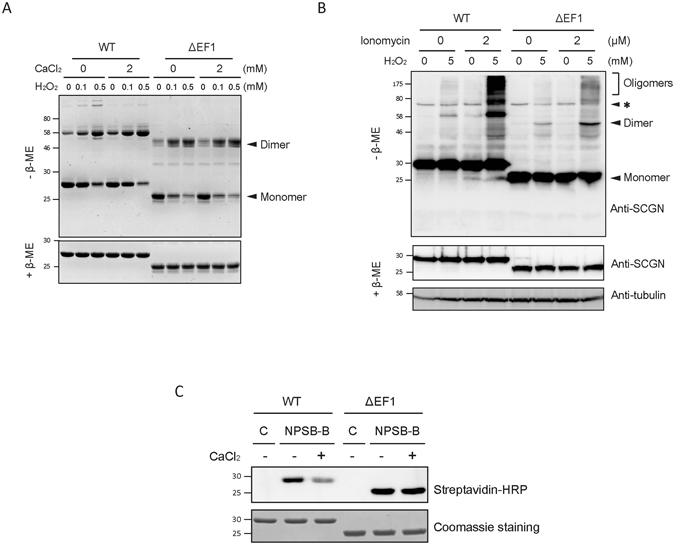
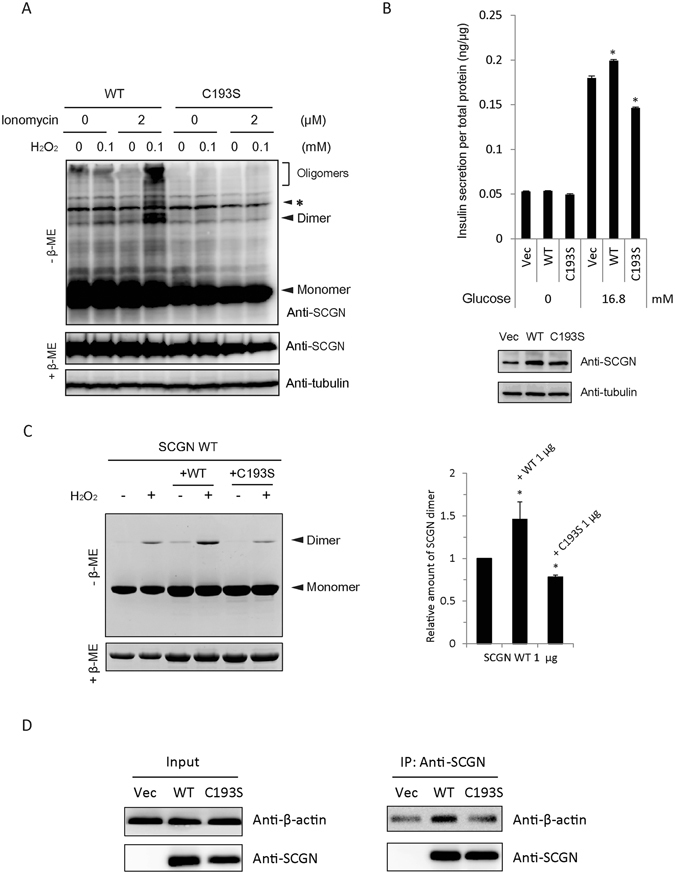
Secretagogin (SCGN), a hexa EF-hand calcium binding protein, plays key roles in insulin secretion in pancreatic β-cells. It is not yet understood how the binding of Ca2+ to human SCGN (hSCGN) promotes secretion. Here we have addressed this question, using mass spectrometry combined with a disulfide searching algorithm DBond. We found that the binding of Ca2+ to hSCGN promotes the dimerization of hSCGN via the formation of a Cys193-Cys193 disulfide bond. Hydrogen/deuterium exchange mass spectrometry (HDX-MS) and molecular dynamics studies revealed that Ca2+ binding to the EF-hands of hSCGN induces significant structural changes that affect the solvent exposure of N-terminal region, and hence the redox sensitivity of the Cys193 residue. These redox sensitivity changes were confirmed using biotinylated methyl-3-nitro-4-(piperidin-1-ylsulfonyl) benzoate (NPSB-B), a chemical probe that specifically labels reactive cysteine sulfhydryls. Furthermore, we found that wild type hSCGN overexpression promotes insulin secretion in pancreatic β cells, while C193S-hSCGN inhibits it. These findings suggest that insulin secretion in pancreatic cells is regulated by Ca2+ and ROS signaling through Ca2+-induced structural changes promoting dimerization of hSCGN.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
