

PRR2, a pseudo-response regulator, promotes salicylic acid and camalexin accumulation during plant immunity
- PMID: 28765536
- PMCID: PMC5539105
- DOI: 10.1038/s41598-017-07535-8
PRR2, a pseudo-response regulator, promotes salicylic acid and camalexin accumulation during plant immunity
Abstract
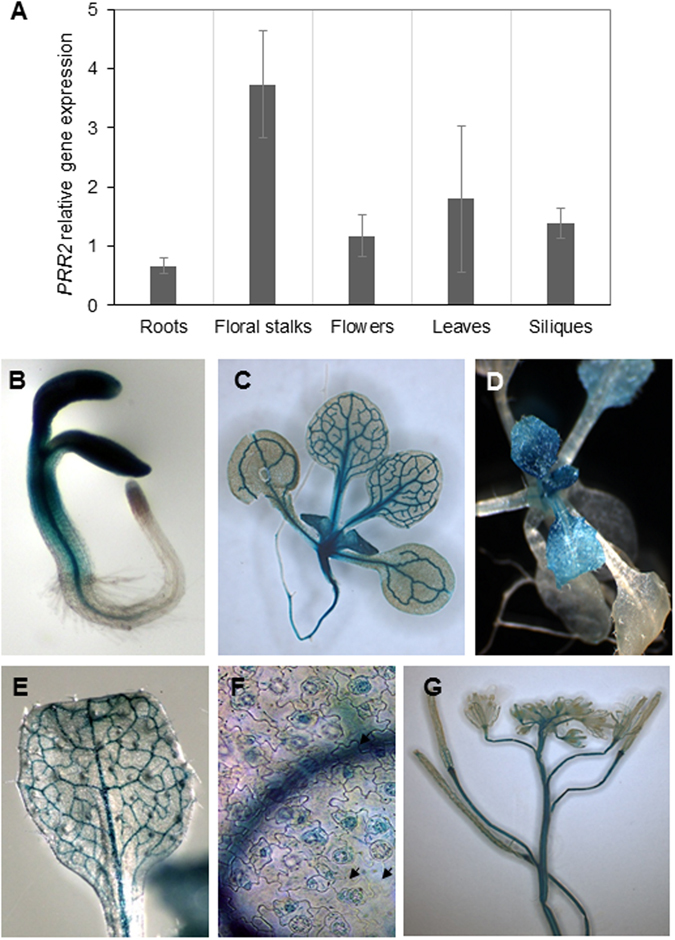
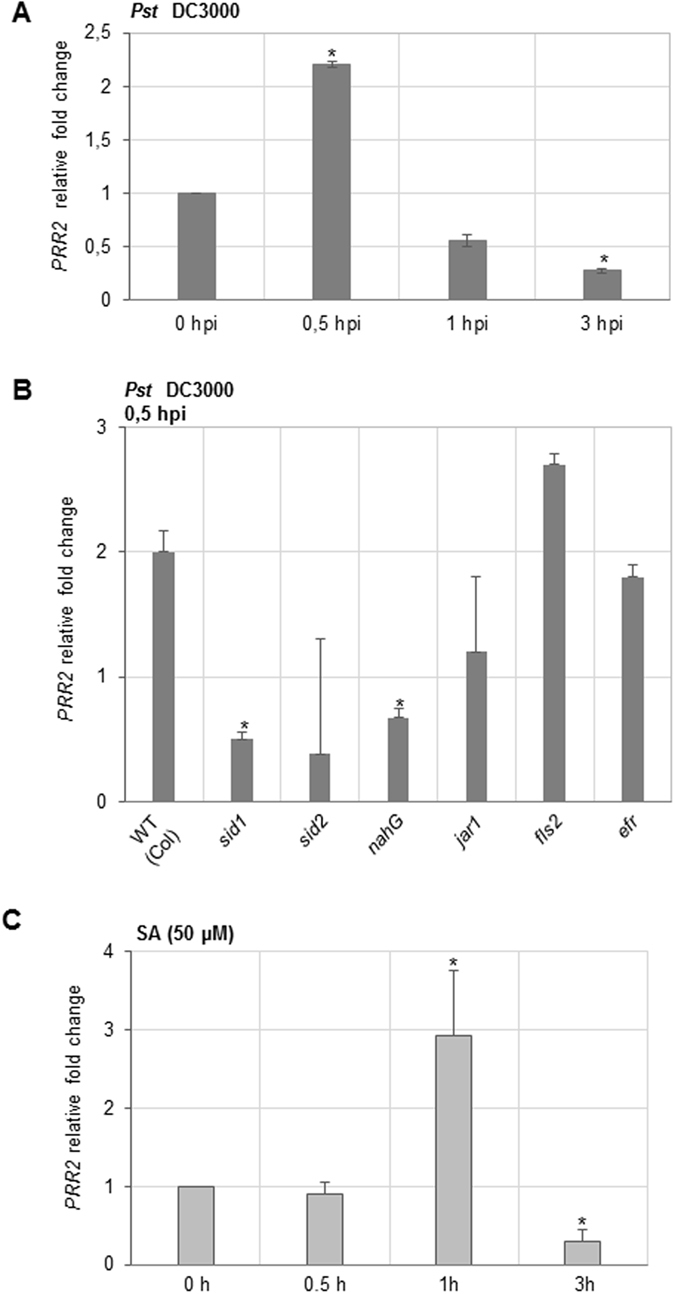
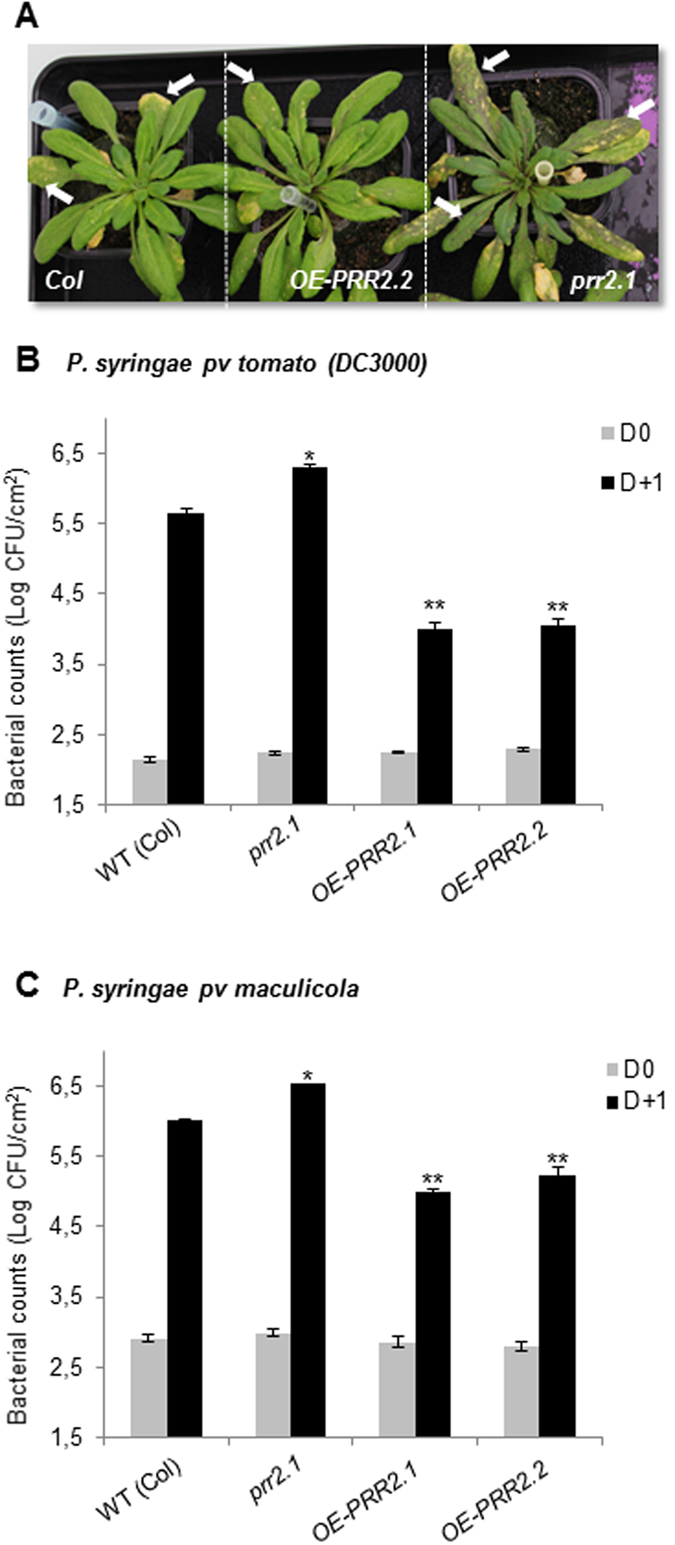
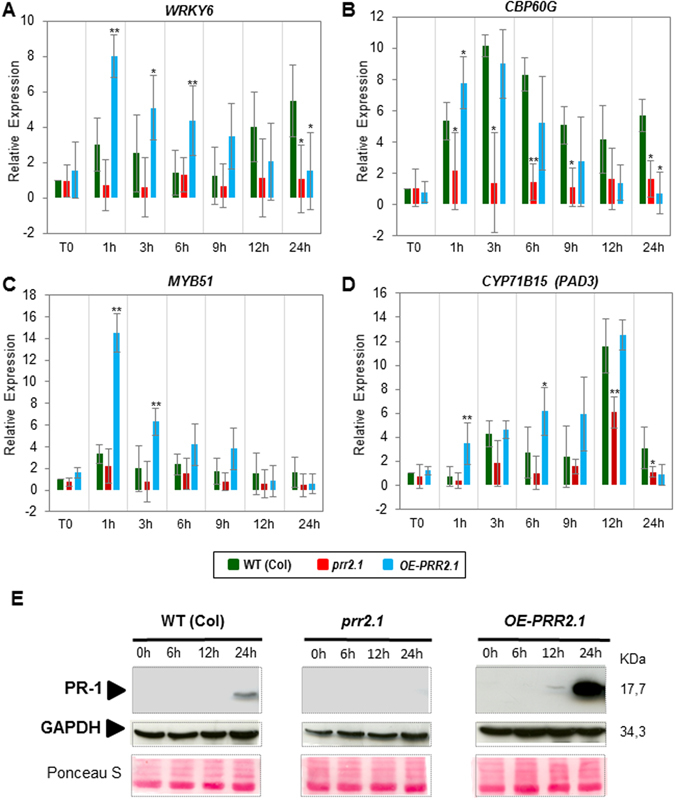
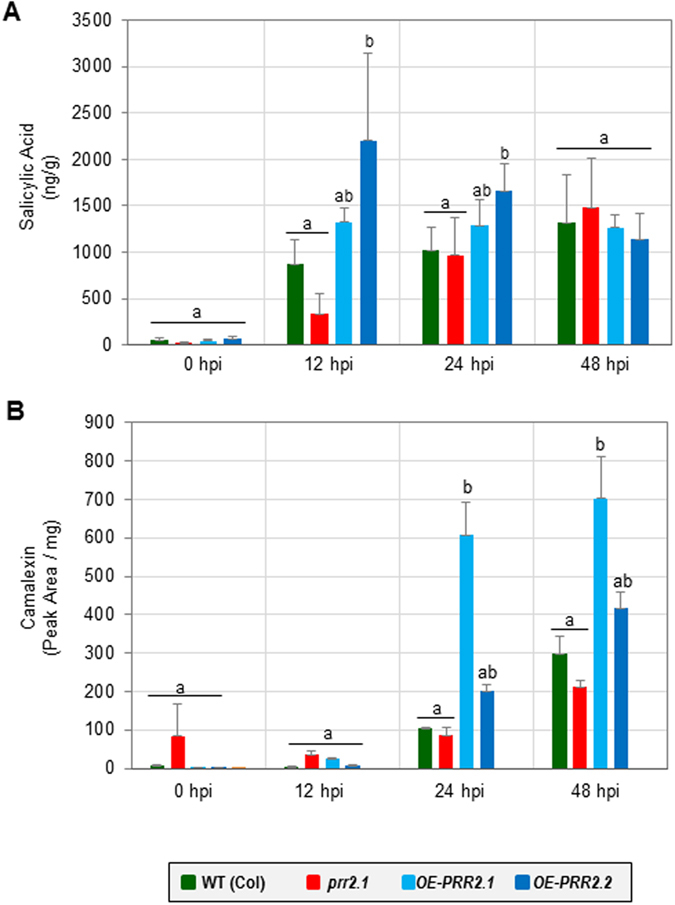
Calcium signalling mediated by Calmodulin (CaM) and calmodulin-like (CML) proteins is critical to plant immunity. CaM and CML regulate a wide range of target proteins and cellular responses. While many CaM-binding proteins have been identified, few have been characterized for their specific role in plant immunity. Here, we report new data on the biological function of a CML-interacting partner, PRR2 (PSEUDO-RESPONSE REGULATOR 2), a plant specific transcription factor. Until now, the physiological relevance of PRR2 remained largely unknown. Using a reverse genetic strategy in A. thaliana, we identified PRR2 as a positive regulator of plant immunity. We propose that PRR2 contributes to salicylic acid (SA)-dependent responses when challenged with the phytopathogenic bacterium Pseudomonas syringae. PRR2 is transcriptionally upregulated by SA and P. syringae, enhances SA biosynthesis and SA signalling responses; e.g. in response to P. syringae, PRR2 induces the production of SA and the accumulation of the defence-related protein PR1. Moreover, PRR2 overexpressing lines exhibit an enhanced production of camalexin, a phytoalexin that confers enhanced resistance against pathogens. Together, these data reveal the importance of PRR2 in plant immune responses against P. syringae and suggest a novel function for this particular plant specific transcription factor in plant physiology.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
