

A CCR2+ myeloid cell niche required for pancreatic β cell growth
- PMID: 28768911
- PMCID: PMC5543911
- DOI: 10.1172/jci.insight.93834
A CCR2+ myeloid cell niche required for pancreatic β cell growth
Abstract
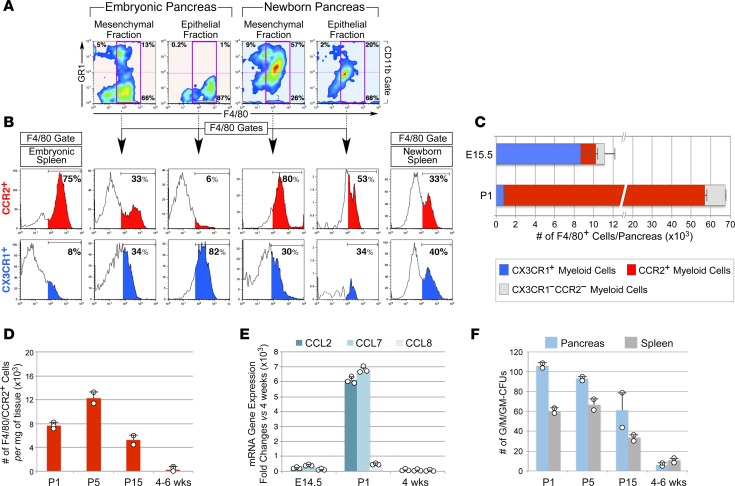
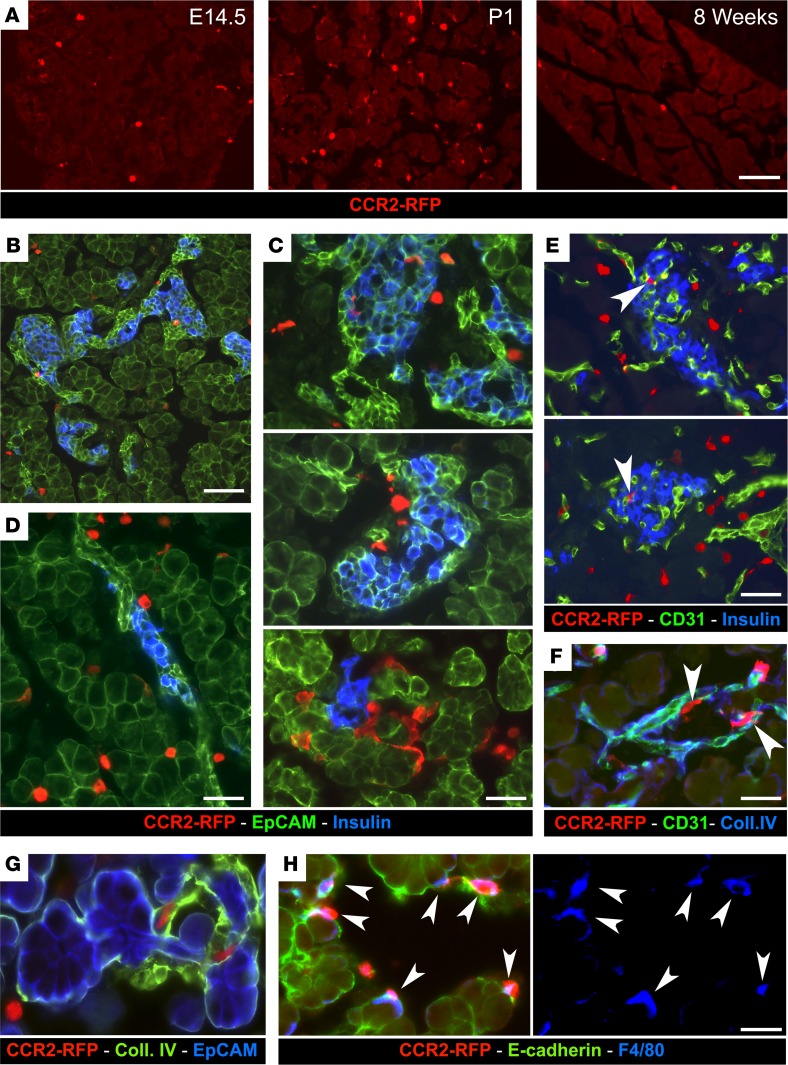
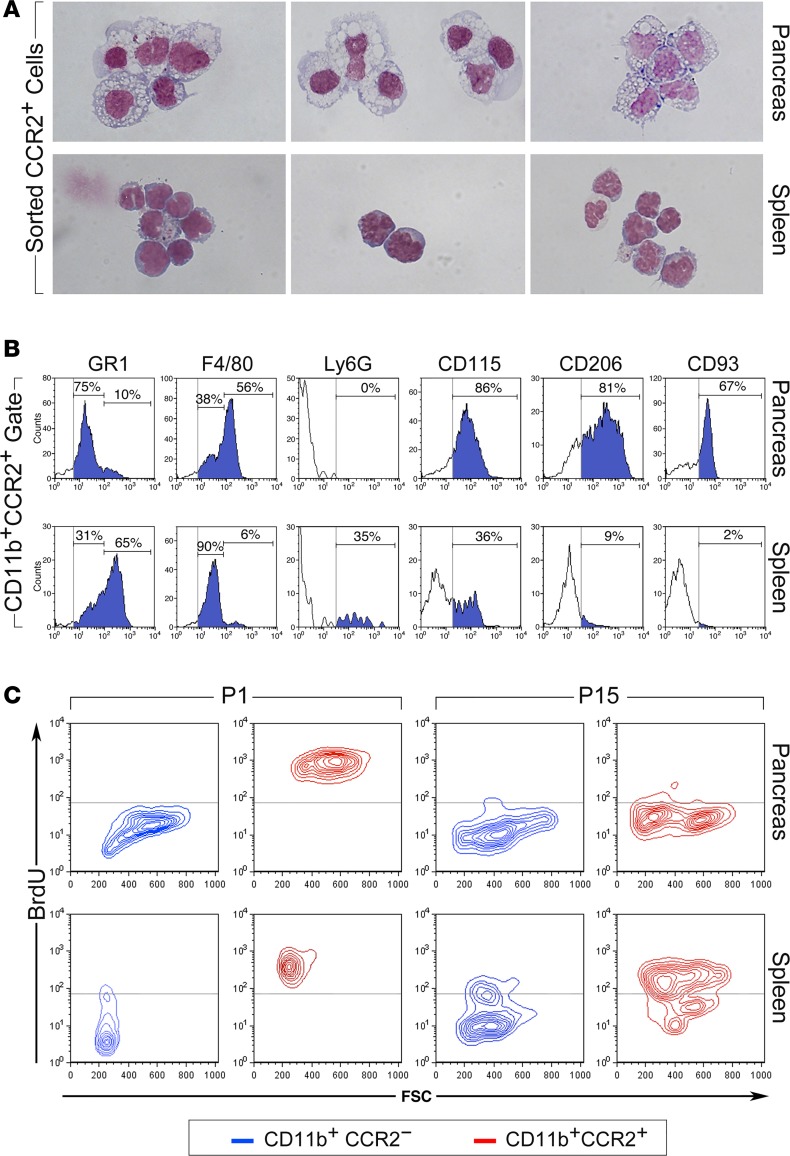
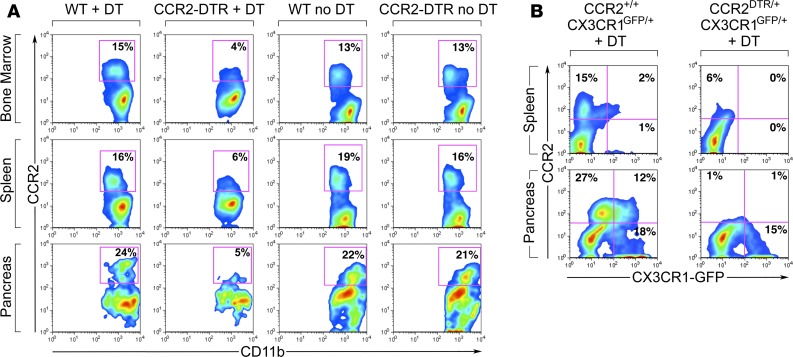
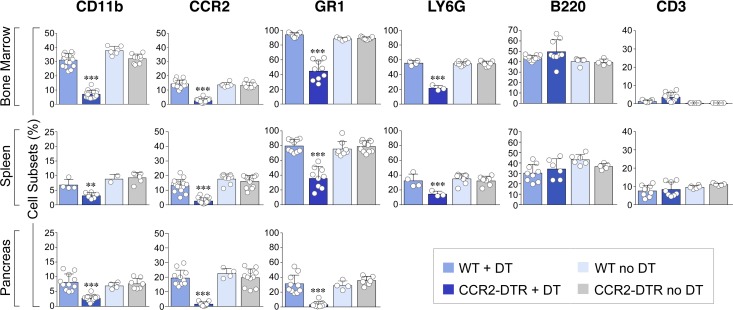
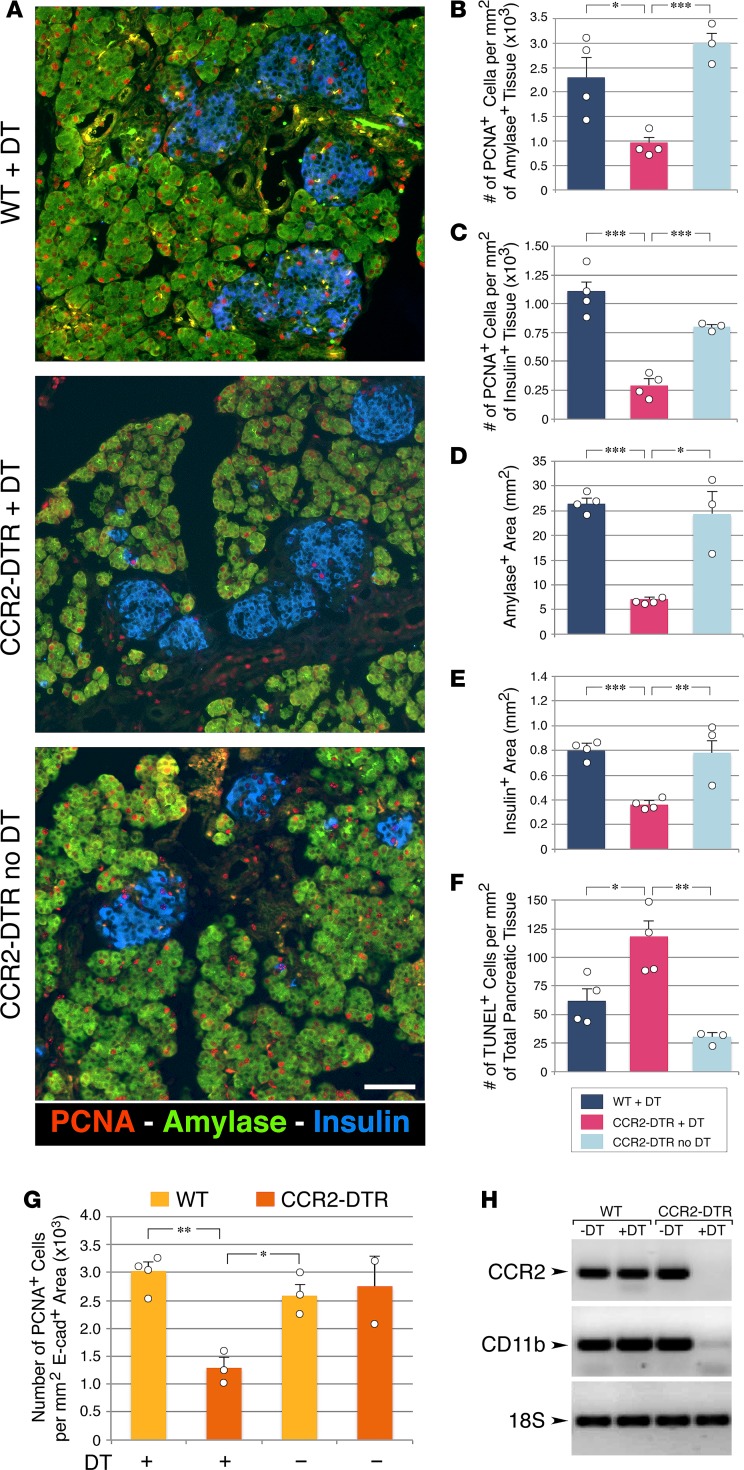
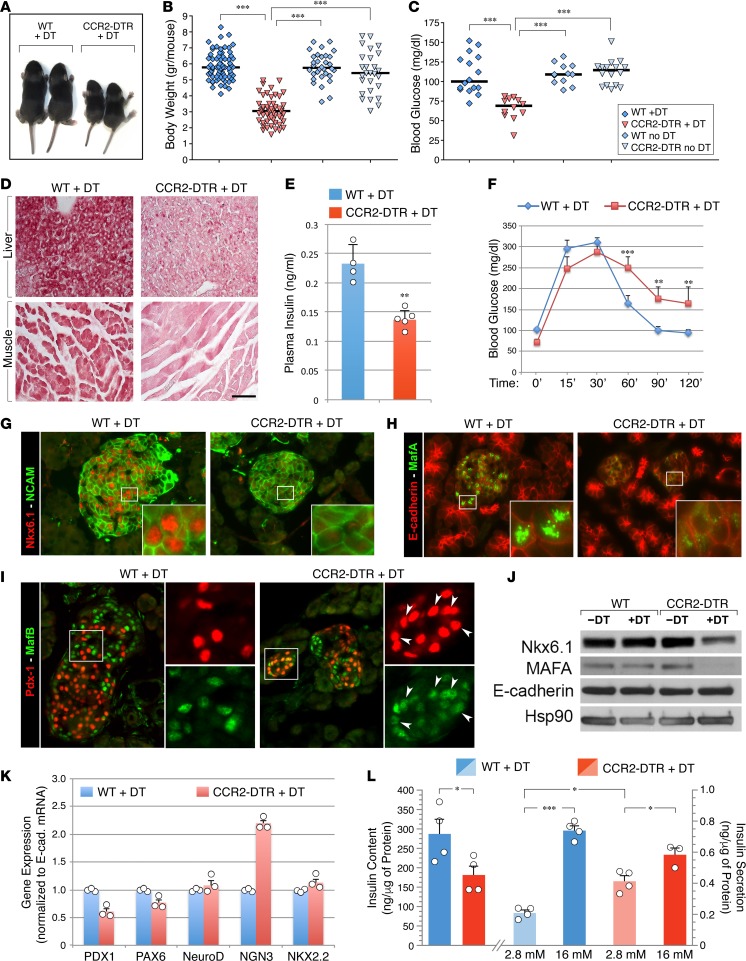
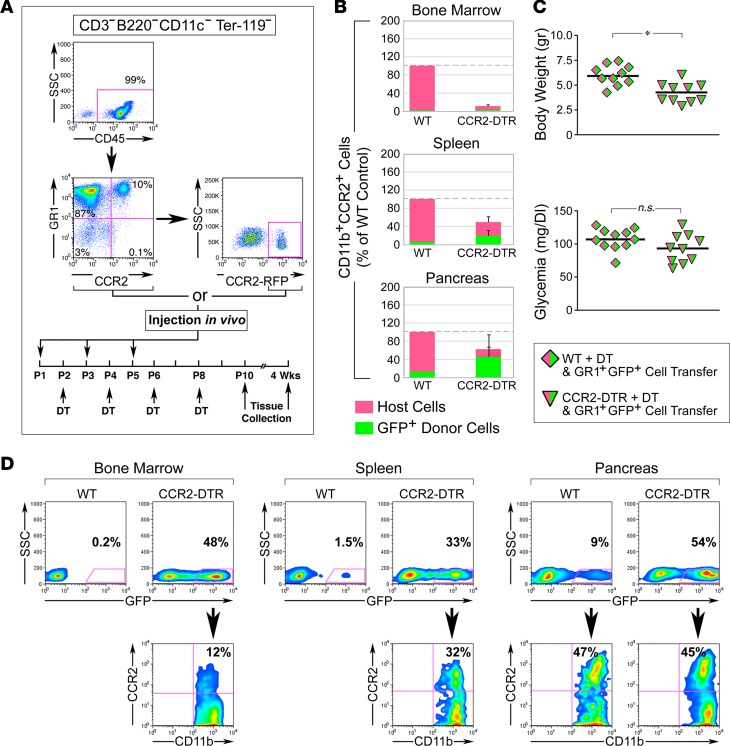
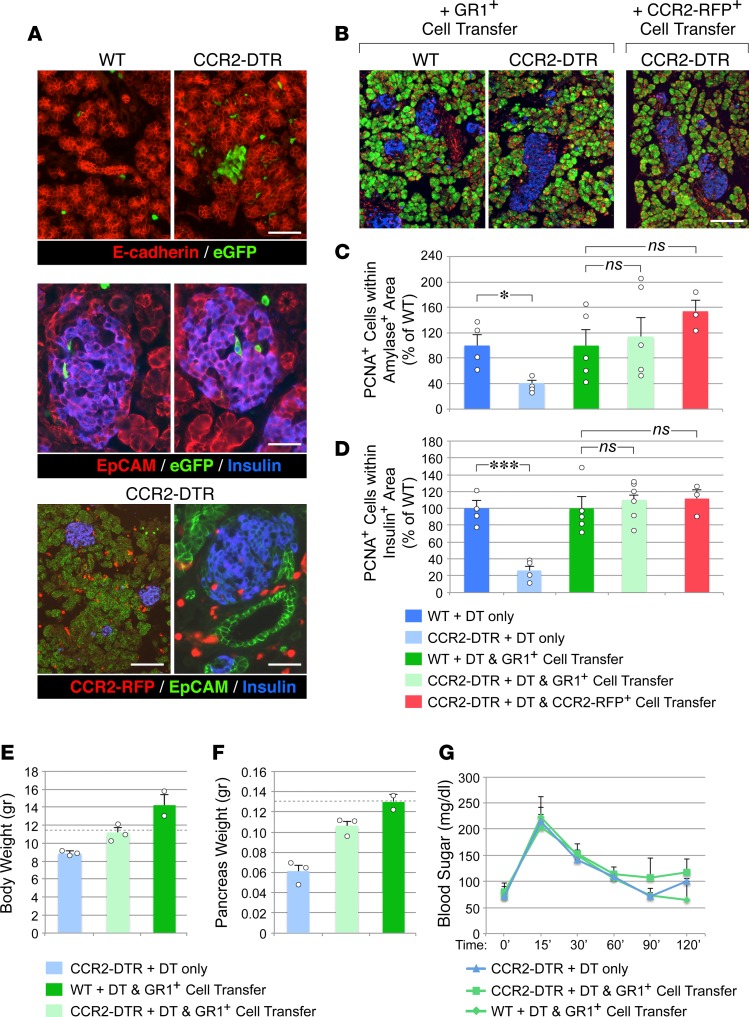
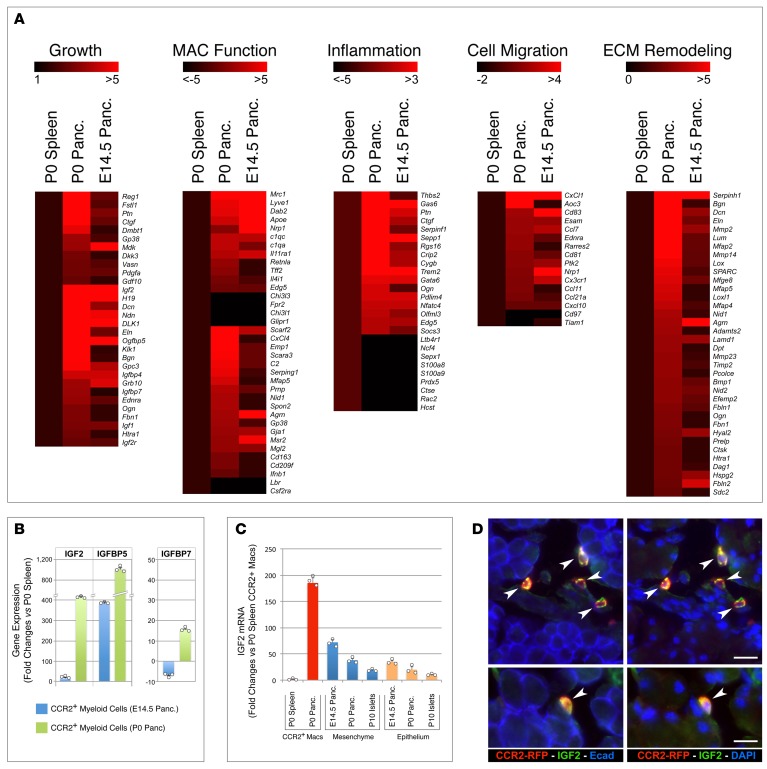
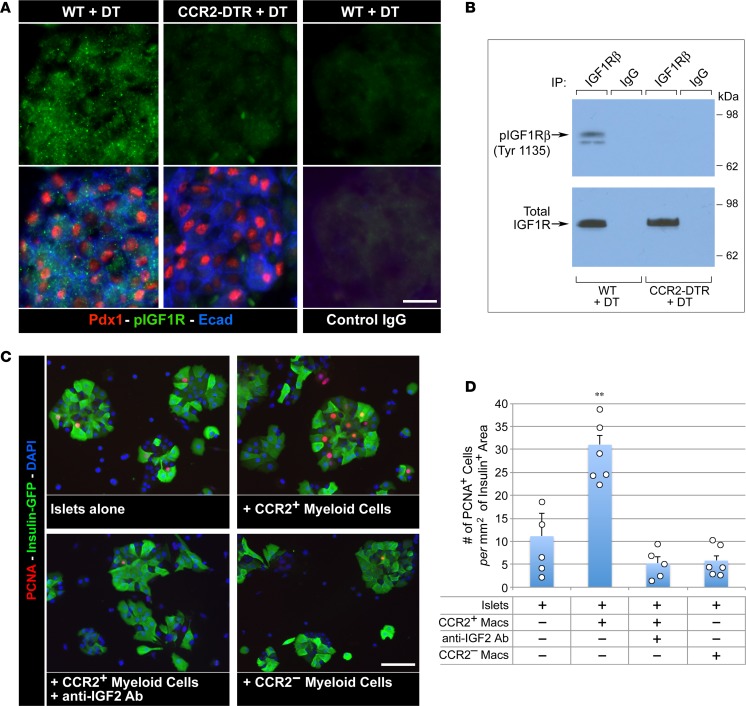
Organ-specific patterns of myeloid cells may contribute tissue-specific growth and/or regenerative potentials. The perinatal stage of pancreas development marks a time characterized by maximal proliferation of pancreatic islets, ensuring the maintenance of glucose homeostasis throughout life. Ontogenically distinct CX3CR1+ and CCR2+ macrophage populations have been reported in the adult pancreas, but their functional contribution to islet cell growth at birth remains unknown. Here, we uncovered a temporally restricted requirement for CCR2+ myeloid cells in the perinatal proliferation of the endocrine pancreatic epithelium. CCR2+ macrophages are transiently enriched over CX3CR1+ subsets in the neonatal pancreas through both local expansion and recruitment of immature precursors. Using CCR2-specific depletion models, we show that loss of this myeloid population leads to a striking reduction in β cell proliferation, dysfunctional islet phenotypes, and glucose intolerance in newborns. Replenishment of pancreatic CCR2+ myeloid compartments by adoptive transfer rescues these defects. Gene profiling identifies pancreatic CCR2+ myeloid cells as a prominent source of IGF2, which contributes to IGF1R-mediated islet proliferation. These findings uncover proproliferative functions of CCR2+ myeloid subsets and identify myeloid-dependent regulation of IGF signaling as a local cue supporting pancreatic proliferation.
Keywords: Development; Endocrinology.
Conflict of interest statement
Figures
Similar articles
-
Neonatal Stroke and TLR1/2 Ligand Recruit Myeloid Cells through the Choroid Plexus in a CX3CR1-CCR2- and Context-Specific Manner.J Neurosci. 2020 May 6;40(19):3849-3861. doi: 10.1523/JNEUROSCI.2149-19.2020. Epub 2020 Apr 8. J Neurosci. 2020. PMID: 32269105 Free PMC article.
-
Chemokine receptors CCR2 and CX3CR1 regulate skin fibrosis in the mouse model of cytokine-induced systemic sclerosis.J Dermatol Sci. 2013 Mar;69(3):250-8. doi: 10.1016/j.jdermsci.2012.10.010. Epub 2012 Oct 24. J Dermatol Sci. 2013. PMID: 23142052
-
Spatio-temporal profile, phenotypic diversity, and fate of recruited monocytes into the post-ischemic brain.J Neuroinflammation. 2016 Nov 4;13(1):285. doi: 10.1186/s12974-016-0750-0. J Neuroinflammation. 2016. PMID: 27814740 Free PMC article.
-
Can pancreatic duct-derived progenitors be a source of islet regeneration?Biochem Biophys Res Commun. 2009 Jun 12;383(4):383-5. doi: 10.1016/j.bbrc.2009.03.114. Epub 2009 Mar 24. Biochem Biophys Res Commun. 2009. PMID: 19324022 Review.
-
Unraveling the role of the ghrelin gene peptides in the endocrine pancreas.J Mol Endocrinol. 2010 Sep;45(3):107-18. doi: 10.1677/JME-10-0019. Epub 2010 Jul 1. J Mol Endocrinol. 2010. PMID: 20595321 Review.
Cited by
-
Islet Regeneration: Endogenous and Exogenous Approaches.Int J Mol Sci. 2021 Mar 24;22(7):3306. doi: 10.3390/ijms22073306. Int J Mol Sci. 2021. PMID: 33804882 Free PMC article. Review.
-
Genetically engineered myeloid cells rebalance the core immune suppression program in metastasis.Cell. 2021 Apr 15;184(8):2033-2052.e21. doi: 10.1016/j.cell.2021.02.048. Epub 2021 Mar 24. Cell. 2021. PMID: 33765443 Free PMC article.
-
Neonatal fungi promote lifelong metabolic health through macrophage-dependent β cell development.Science. 2025 Mar 7;387(6738):eadn0953. doi: 10.1126/science.adn0953. Epub 2025 Mar 7. Science. 2025. PMID: 40048508
-
Understanding the Biology of Self-Renewing Macrophages.Cells. 2018 Aug 9;7(8):103. doi: 10.3390/cells7080103. Cells. 2018. PMID: 30096862 Free PMC article. Review.
-
Pericytes modulate islet immune cells and insulin secretion through Interleukin-33 production in mice.Front Endocrinol (Lausanne). 2023 Mar 9;14:1142988. doi: 10.3389/fendo.2023.1142988. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 36967785 Free PMC article.
References
-
- Wiktor-Jedrzejczak W, et al. Correction by CSF-1 of defects in the osteopetrotic op/op mouse suggests local, developmental, and humoral requirements for this growth factor. Exp Hematol. 1991;19(10):1049–1054. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
