

Connexin 43 is required for the maintenance of mitochondrial integrity in brown adipose tissue
- PMID: 28769076
- PMCID: PMC5540980
- DOI: 10.1038/s41598-017-07658-y
Connexin 43 is required for the maintenance of mitochondrial integrity in brown adipose tissue
Abstract
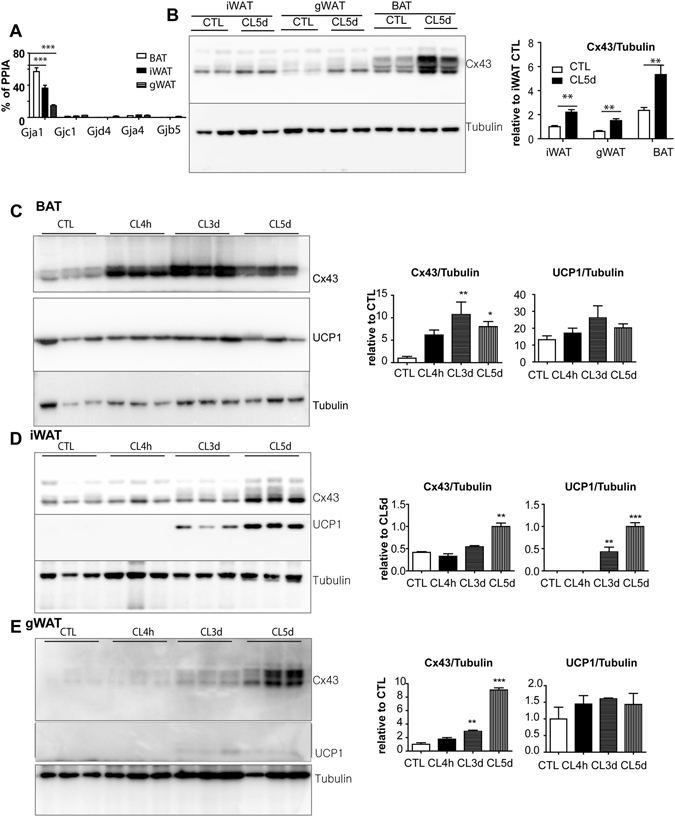
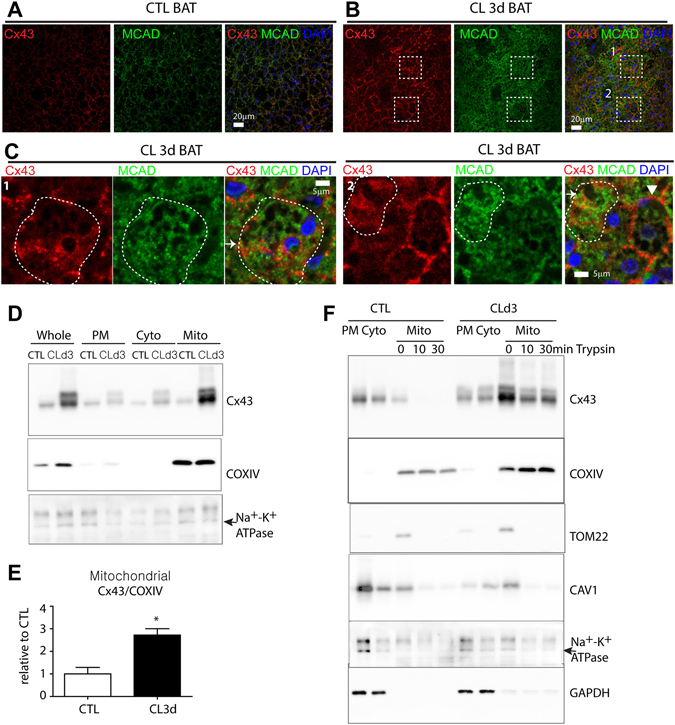
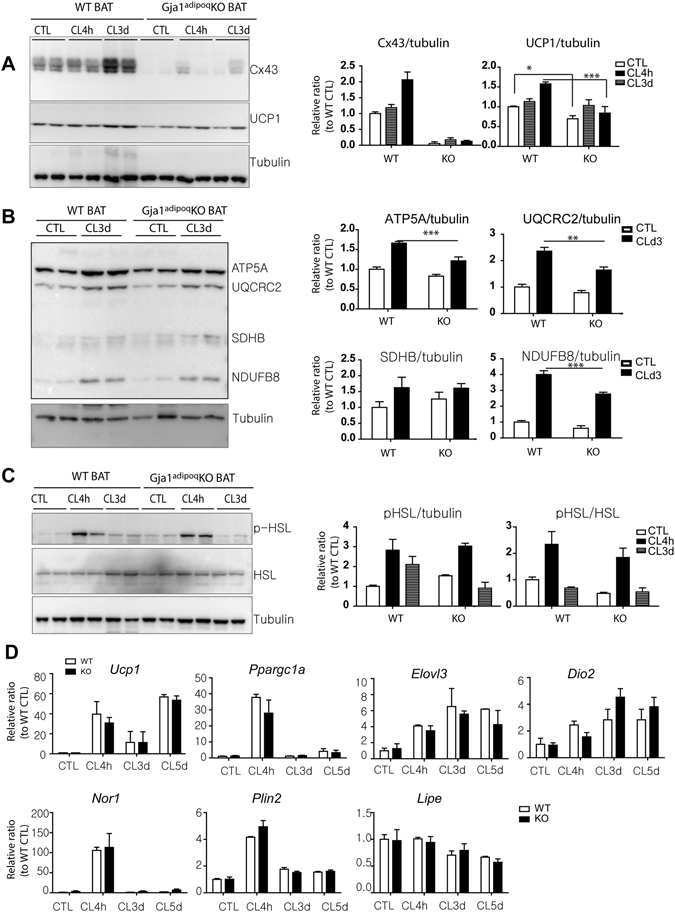
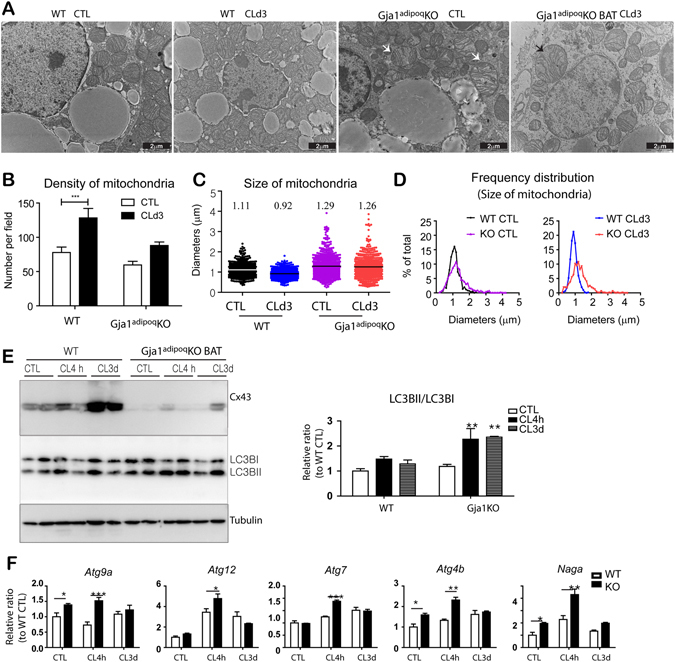
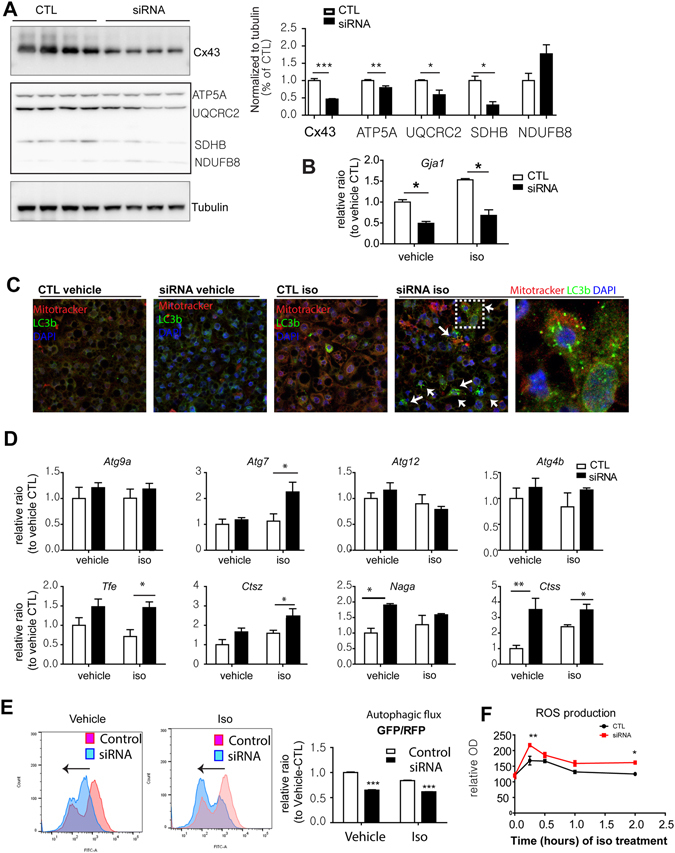
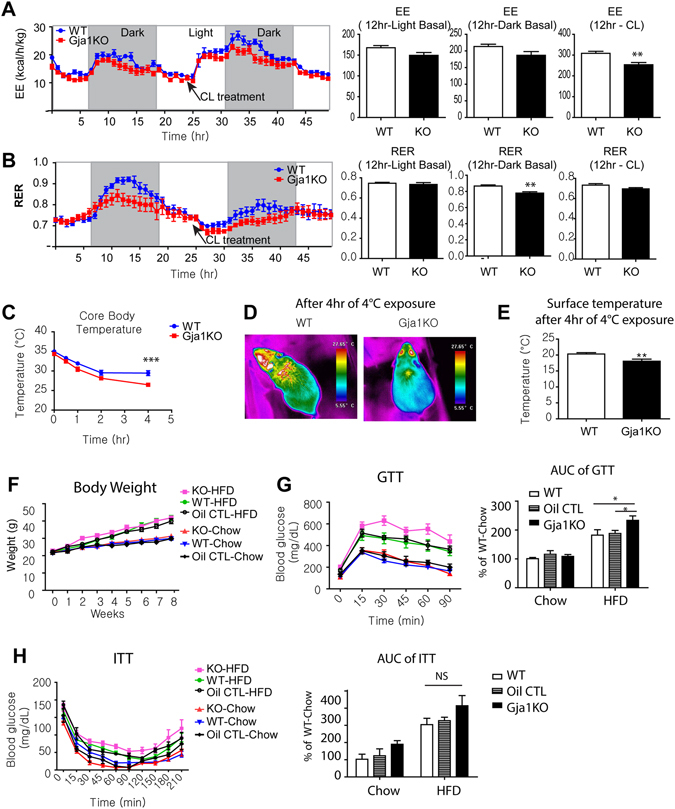
We investigated the role of connexin 43 (Cx43) in maintaining the integrity of mitochondria in brown adipose tissue (BAT). The functional effects of Cx43 were evaluated using inducible, adipocyte-specific Cx43 knockout in mice (Gja1 adipoq KO) and by overexpression and knockdown of Cx43 in cultured adipocytes. Mitochondrial morphology was evaluated by electron microscopy and mitochondrial function and autophagy were assessed by immunoblotting, immunohistochemistry, and qPCR. The metabolic effects of adipocyte-specific knockout of Cx43 were assessed during cold stress and following high fat diet feeding. Cx43 expression was higher in BAT compared to white adipose tissue. Treatment with the β3-adrenergic receptor agonist CL316,243 increased Cx43 expression and mitochondrial localization. Gja1 adipoq KO mice reduced mitochondrial density and increased the presence of damaged mitochondria in BAT. Moreover, metabolic activation with CL316,243 further reduced mitochondrial integrity and upregulated autophagy in the BAT of Gja1 adipoq KO mice. Inhibition of Cx43 in cultured adipocytes increased the generation of reactive oxygen species and induction of autophagy during β-adrenergic stimulation. Gja1 adipoq KO mice were cold intolerant, expended less energy in response to β3-adrenergic receptor activation, and were more insulin resistant after a high-fat diet challenge. Collectively, our data demonstrate that Cx43 is required for maintaining the mitochondrial integrity and metabolic activity of BAT.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
