

Functions and regulation of the Polo-like kinase Cdc5 in the absence and presence of DNA damage
- PMID: 28770345
- PMCID: PMC6249032
- DOI: 10.1007/s00294-017-0727-2
Functions and regulation of the Polo-like kinase Cdc5 in the absence and presence of DNA damage
Abstract
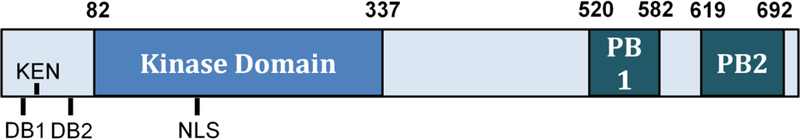
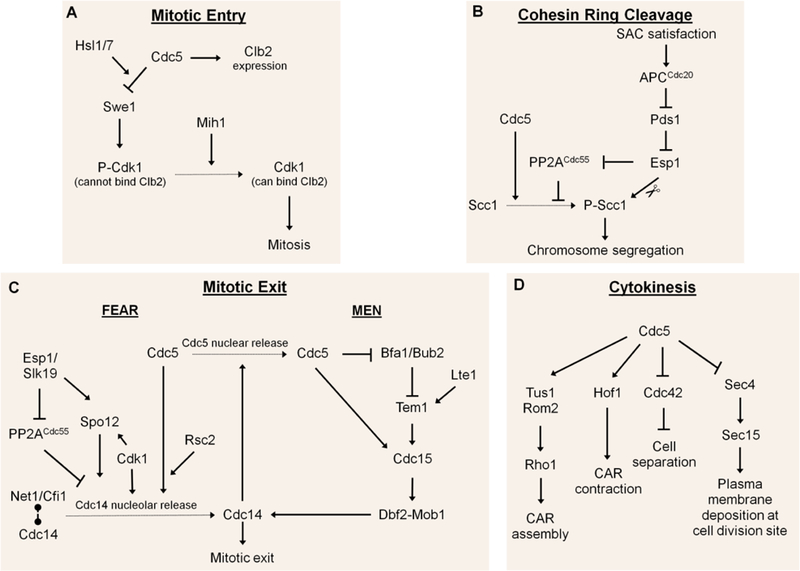
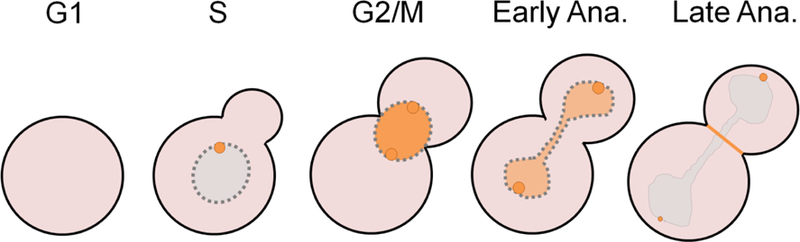
Polo-like kinases are essential cell cycle regulators that are conserved from yeast to humans. Unlike higher eukaryotes, who express multiple Polo-like kinase family members that perform many important functions, budding yeast express only a single Polo-like kinase, Cdc5, which is the homolog of mammalian cell cycle master regulator Polo-like kinase 1. Cdc5 is a fascinating multifaceted protein that is programmed to target its many substrates in a timely, sequential manner to ensure proper cell cycle progression. Over the years, many lessons about Polo-like kinase 1 have been learned by studying Cdc5 in budding yeast. Cdc5 has been well documented in regulating mitotic entry, chromosome segregation, mitotic exit, and cytokinesis. Cdc5 also plays important roles during cell division after DNA damage. Here, we briefly review the many functions of Cdc5 and its regulation in the absence and presence of DNA damage.
Keywords: Cdc5; Cell cycle; DNA damage; Mitosis; Plk1; Polo-like kinase 1.
Figures
References
-
- Alexandru G, Uhlmann F, Mechtler K, Poupart MA, Nasmyth K (2001) Phosphorylation of the cohesin subunit Scc1 by Polo/Cdc5 kinase regulates sister chromatid separation in yeast Cell 105:459–472 - PubMed
-
- Archambault V, Glover DM (2009) Polo-like kinases: conservation and divergence in their functions and regulation Nat Rev Mol Cell Biol 10:265–275 10.1038/nrm2653 - DOI - PubMed
-
- Arnold L, Hockner S, Seufert W (2015) Insights into the cellular mechanism of the yeast ubiquitin ligase APC/C-Cdh1 from the analysis of in vivo degrons Mol Biol Cell 26:843–858 10.1091/mbc.E14-09-1342 - DOI - PMC - PubMed
-
- Asano S et al. (2005) Concerted mechanism of Swe1/Wee1 regulation by multiple kinases in budding yeast EMBO J 24:2194–2204 10.1038/sj.emboj.7600683 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
