

Identification and characterization of a novel botulinum neurotoxin
- PMID: 28770820
- PMCID: PMC5543303
- DOI: 10.1038/ncomms14130
Identification and characterization of a novel botulinum neurotoxin
Abstract
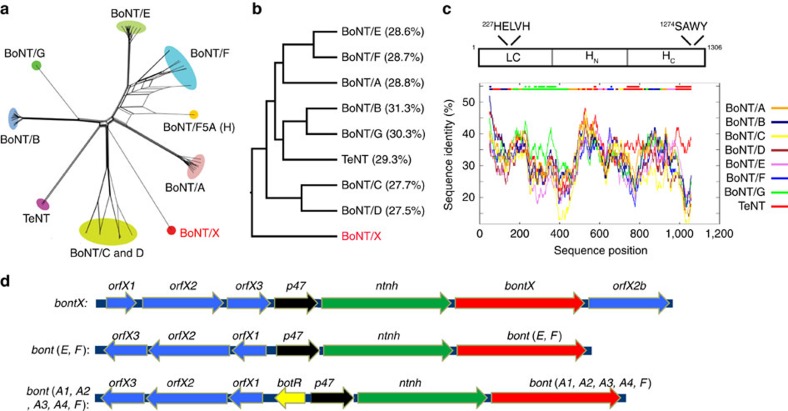
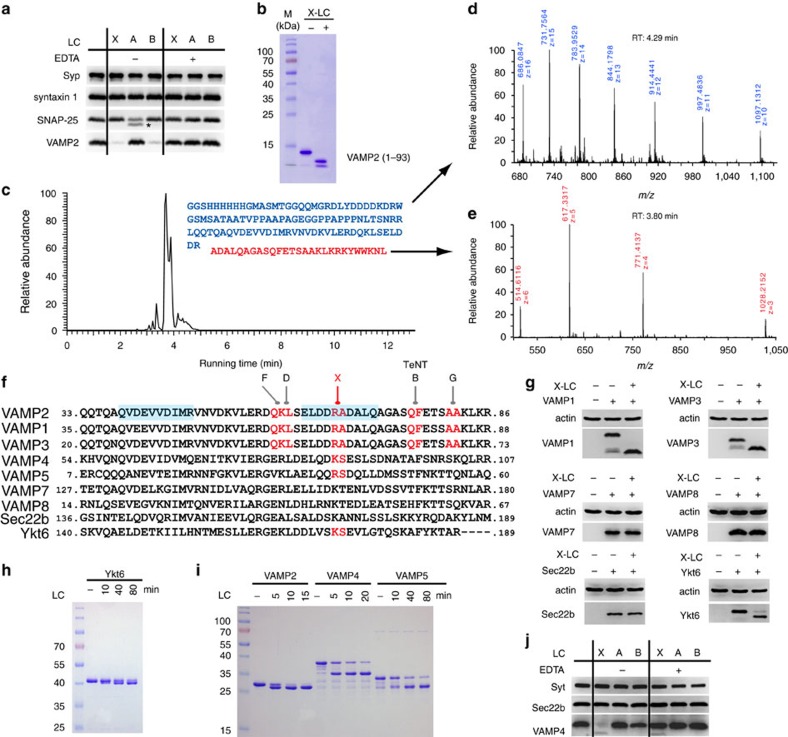
Botulinum neurotoxins are known to have seven serotypes (BoNT/A-G). Here we report a new BoNT serotype, tentatively named BoNT/X, which has the lowest sequence identity with other BoNTs and is not recognized by antisera against known BoNTs. Similar to BoNT/B/D/F/G, BoNT/X cleaves vesicle-associated membrane proteins (VAMP) 1, 2 and 3, but at a novel site (Arg66-Ala67 in VAMP2). Remarkably, BoNT/X is the only toxin that also cleaves non-canonical substrates VAMP4, VAMP5 and Ykt6. To validate its activity, a small amount of full-length BoNT/X was assembled by linking two non-toxic fragments using a transpeptidase (sortase). Assembled BoNT/X cleaves VAMP2 and VAMP4 in cultured neurons and causes flaccid paralysis in mice. Thus, BoNT/X is a novel BoNT with a unique substrate profile. Its discovery posts a challenge to develop effective countermeasures, provides a novel tool for studying intracellular membrane trafficking, and presents a new potential therapeutic toxin for modulating secretions in cells.
Conflict of interest statement
Boston Children’s Hospital has filed a provisional patent application (U.S. 62/360,239) for medical use of BoNT/X, with P.S., S.Z. and M.D. as inventors.
Figures
References
-
- Arnon S. S. et al. Botulinum toxin as a biological weapon: medical and public health management. JAMA 285, 1059–1070 (2001). - PubMed
-
- Johnson E. A. Clostridial toxins as therapeutic agents: benefits of nature’s most toxic proteins. Annu. Rev. Microbiol. 53, 551–575 (1999). - PubMed
-
- Montecucco C. & Molgo J. Botulinal neurotoxins: revival of an old killer. Curr. Opin. Pharmacol. 5, 274–279 (2005). - PubMed
-
- Schiavo G., Matteoli M. & Montecucco C. Neurotoxins affecting neuroexocytosis. Physiol. Rev. 80, 717–766 (2000). - PubMed
-
- Montal M. Botulinum neurotoxin: a marvel of protein design. Annu. Rev. Biochem. 79, 591–617 (2010). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
