

FGFR1 is critical for the anti-endothelial mesenchymal transition effect of N-acetyl-seryl-aspartyl-lysyl-proline via induction of the MAP4K4 pathway
- PMID: 28771231
- PMCID: PMC5596544
- DOI: 10.1038/cddis.2017.353
FGFR1 is critical for the anti-endothelial mesenchymal transition effect of N-acetyl-seryl-aspartyl-lysyl-proline via induction of the MAP4K4 pathway
Abstract
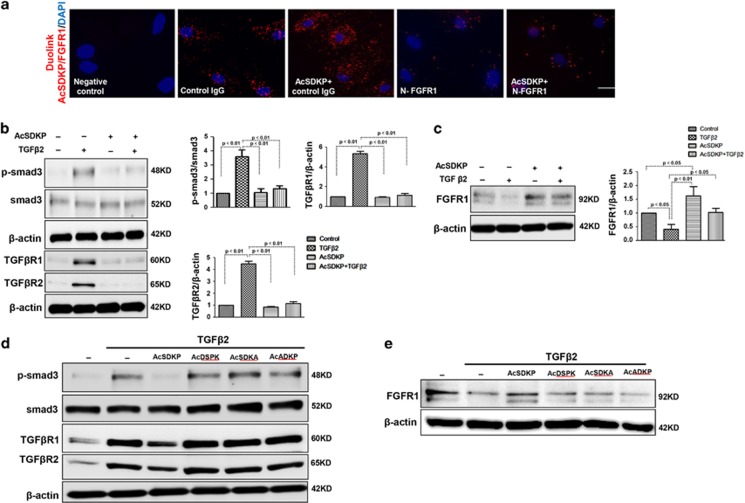
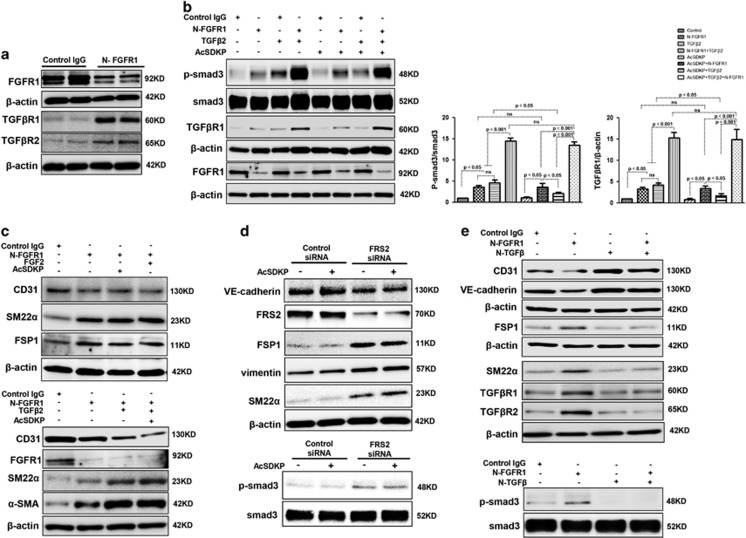
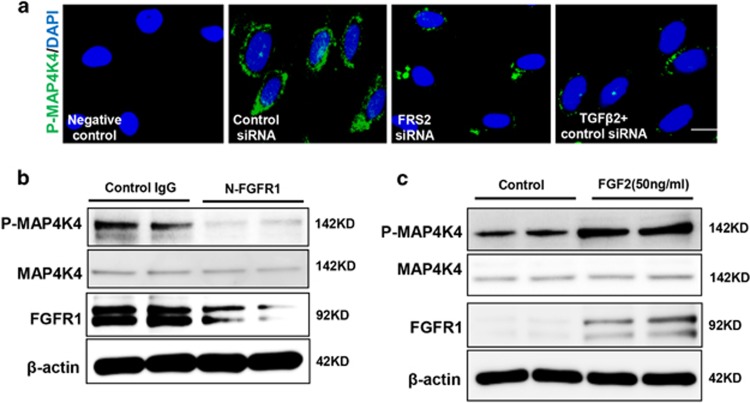
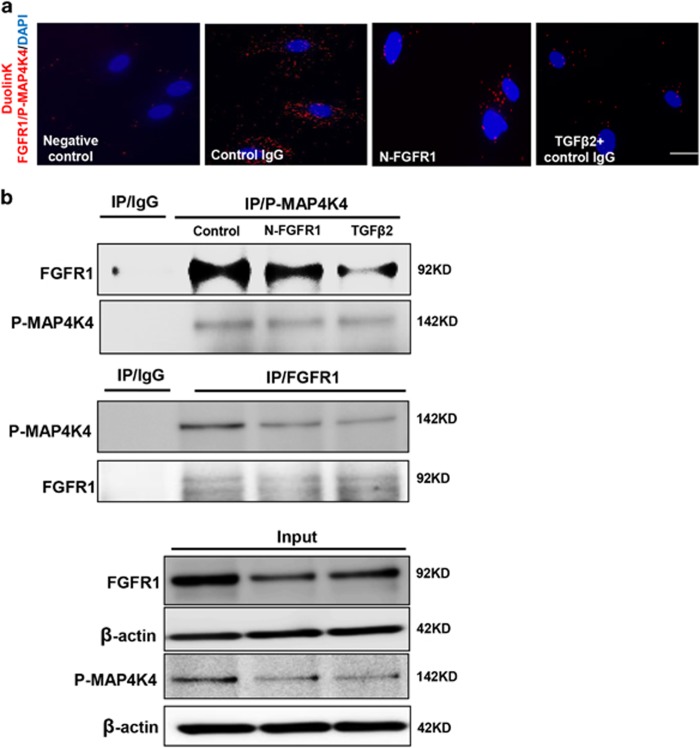
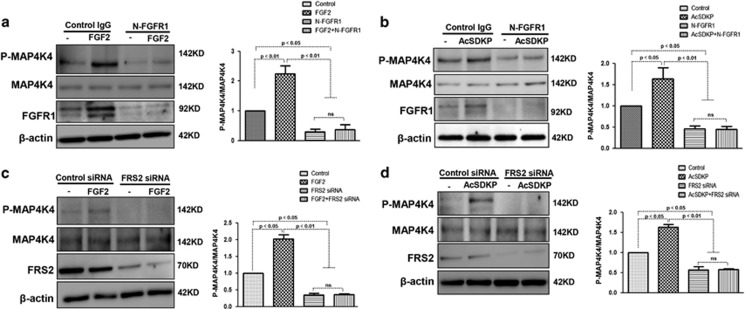
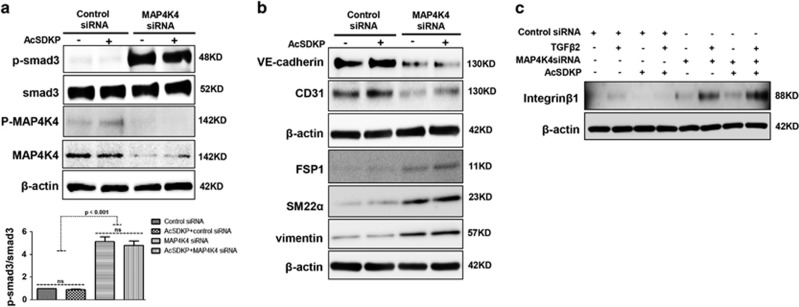
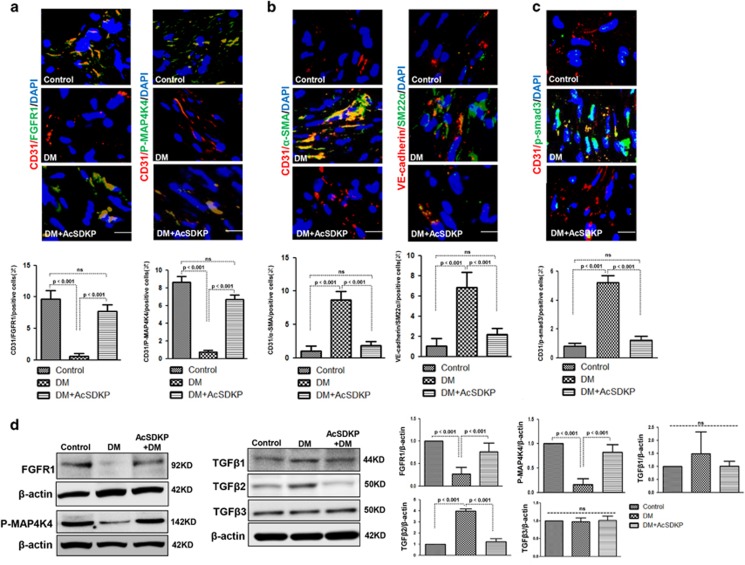
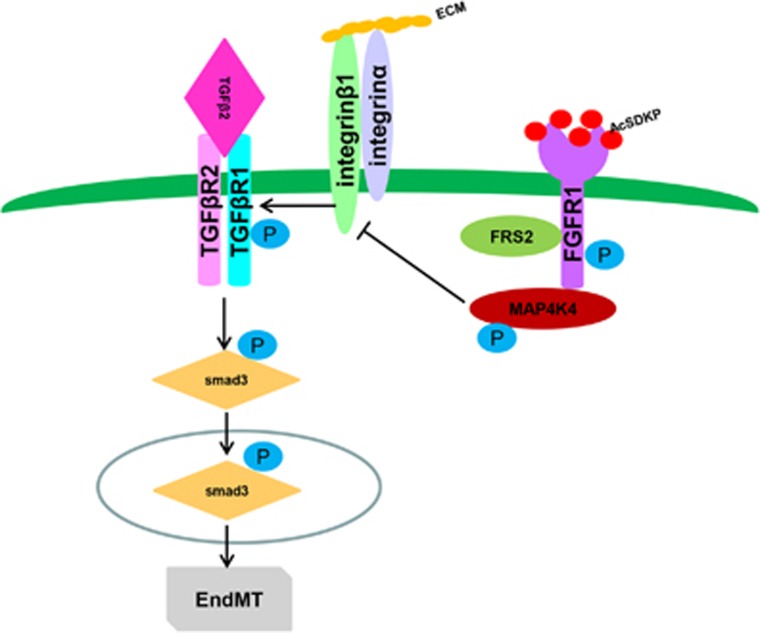
Endothelial-to-mesenchymal transition (EndMT) has been shown to contribute to organ fibrogenesis, and we have reported that the anti-EndMT effect of N-acetyl-seryl-aspartyl-lysyl-proline (AcSDKP) is associated with restoring expression of diabetes-suppressed fibroblast growth factor receptor (FGFR), the key anti-EndMT molecule. FGFR1 is the key inhibitor of EndMT via the suppression of the transforming growth factor β (TGFβ) signaling pathway, and mitogen-activated protein kinase kinase kinase kinase 4 (MAP4K4) inhibits integrin β1, a key factor in activating TGFβ signaling and EndMT. Here, we showed that the close proximity between AcSDKP and FGFR1 was essential for the suppression of TGFβ/smad signaling and EndMT associated with MAP4K4 phosphorylation (P-MAP4K4) in endothelial cells. In cultured human dermal microvascular endothelial cells (HMVECs), the anti-EndMT and anti-TGFβ/smad effects of AcSDKP were lost following treatment with a neutralizing FGFR1 antibody (N-FGFR1) or transfection of FRS2 siRNA. The physical interaction between FGFR1 and P-MAP4K4 in HMVECs was confirmed by proximity ligation analysis and an immunoprecipitation assay. AcSDKP induced P-MAP4K4 in HMVECs, which was significantly inhibited by treatment with either N-FGFR1 or FRS2 siRNA. Furthermore, MAP4K4 knockdown using specific siRNAs induced smad3 phosphorylation and EndMT in HMVECs, which was not suppressed by AcSDKP. Streptozotocin-induced diabetic CD-1 mice exhibited suppression of both FGFR1 and P-MAP4K4 expression levels associated with the induction of TGFβ/smad3 signaling and EndMT in their hearts and kidneys; those were restored by AcSDKP treatment. These data demonstrate that the AcSDKP-FGFR1-MAP4K4 axis has an important role in combating EndMT-associated fibrotic disorders.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- He J, Xu Y, Koya D, Kanasaki K. Role of the endothelial-to-mesenchymal transition in renal fibrosis of chronic kidney disease. Clin Exp Nephrol 2013; 17: 488–497. - PubMed
-
- Zeisberg EM, Tarnavski O, Zeisberg M, Dorfman AL, McMullen JR, Gustafsson E et al. Endothelial-to-mesenchymal transition contributes to cardiac fibrosis. Nat Med 2007; 13: 952–961. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
