

Deconstructing isolation-by-distance: The genomic consequences of limited dispersal
- PMID: 28771477
- PMCID: PMC5542401
- DOI: 10.1371/journal.pgen.1006911
Deconstructing isolation-by-distance: The genomic consequences of limited dispersal
Abstract
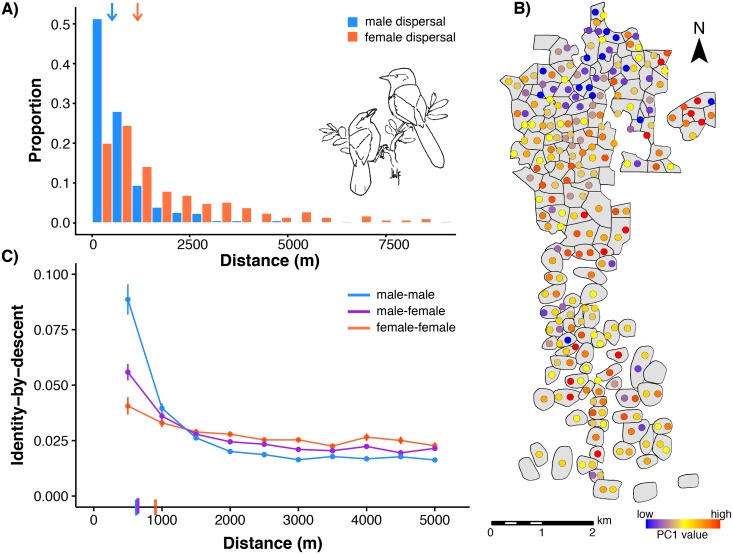
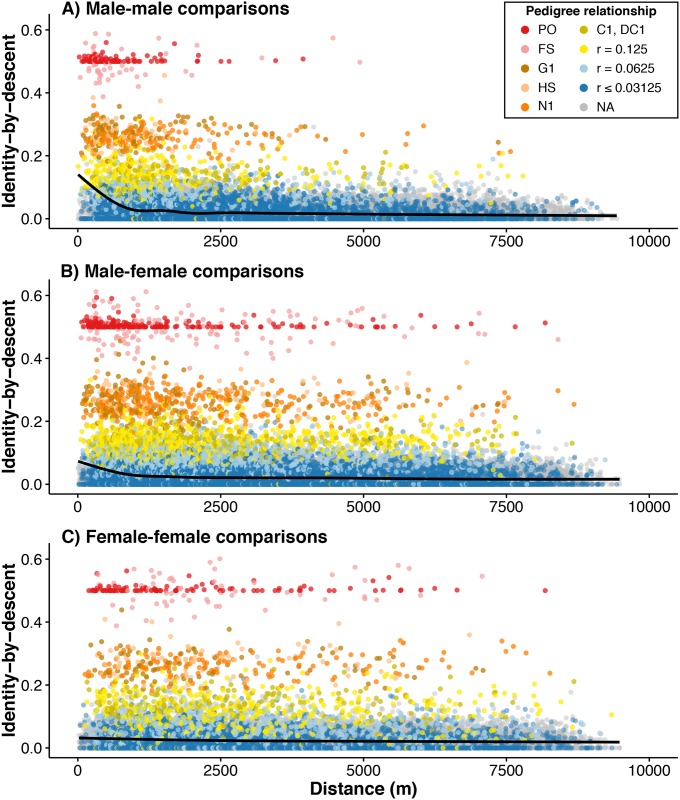
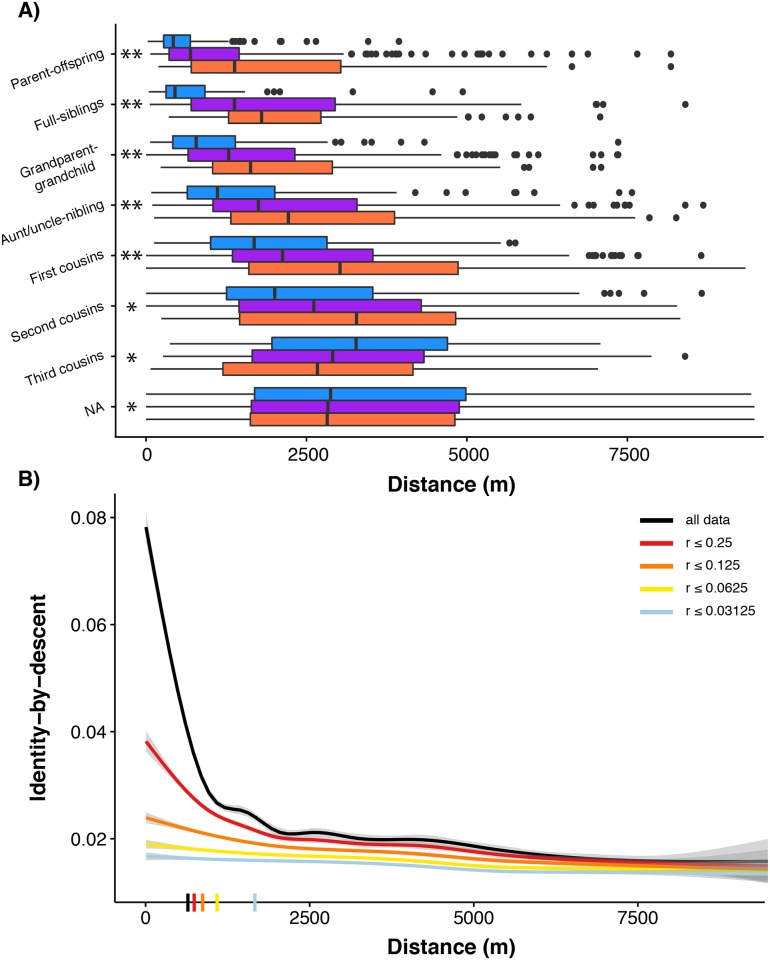
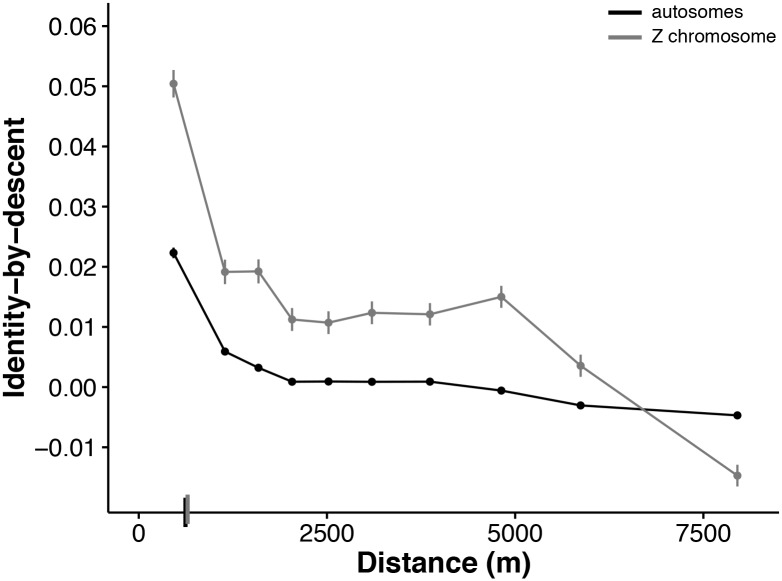
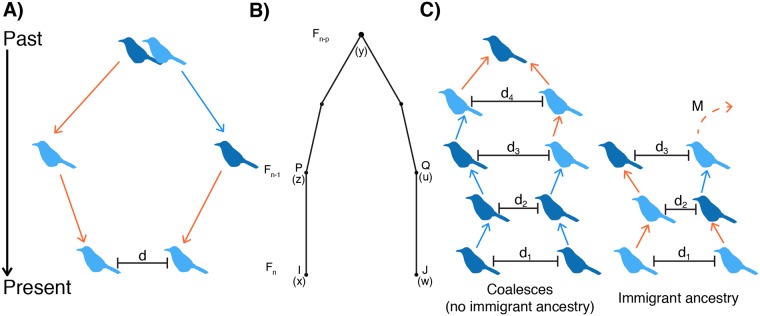
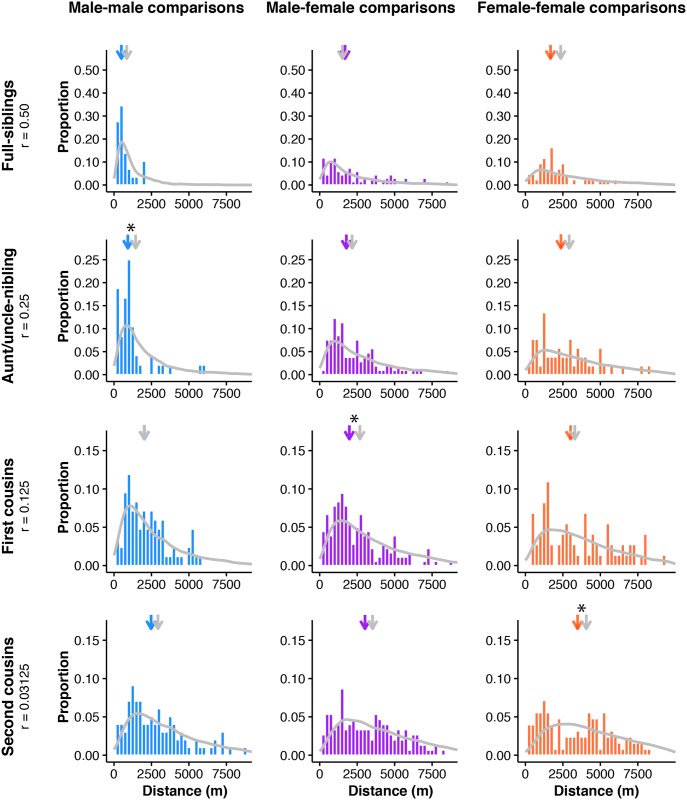
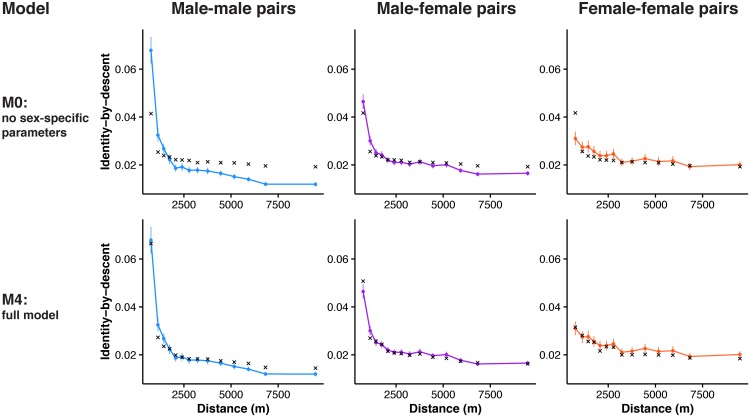
Geographically limited dispersal can shape genetic population structure and result in a correlation between genetic and geographic distance, commonly called isolation-by-distance. Despite the prevalence of isolation-by-distance in nature, to date few studies have empirically demonstrated the processes that generate this pattern, largely because few populations have direct measures of individual dispersal and pedigree information. Intensive, long-term demographic studies and exhaustive genomic surveys in the Florida Scrub-Jay (Aphelocoma coerulescens) provide an excellent opportunity to investigate the influence of dispersal on genetic structure. Here, we used a panel of genome-wide SNPs and extensive pedigree information to explore the role of limited dispersal in shaping patterns of isolation-by-distance in both sexes, and at an exceedingly fine spatial scale (within ~10 km). Isolation-by-distance patterns were stronger in male-male and male-female comparisons than in female-female comparisons, consistent with observed differences in dispersal propensity between the sexes. Using the pedigree, we demonstrated how various genealogical relationships contribute to fine-scale isolation-by-distance. Simulations using field-observed distributions of male and female natal dispersal distances showed good agreement with the distribution of geographic distances between breeding individuals of different pedigree relationship classes. Furthermore, we built coalescent simulations parameterized by the observed dispersal curve, population density, and immigration rate, and showed how incorporating these extensions to Malécot's theory of isolation-by-distance allows us to accurately reconstruct observed sex-specific isolation-by-distance patterns in autosomal and Z-linked SNPs. Therefore, patterns of fine-scale isolation-by-distance in the Florida Scrub-Jay can be well understood as a result of limited dispersal over contemporary timescales.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Clobert J, Danchin E, Dhont AA, Nichols JD. Dispersal. Oxford, UK: Oxford University Press; 2001.
-
- Coulon A, Fitzpatrick JW, Bowman R, Lovette IJ. Mind the gap: genetic distance increases with habitat gap size in Florida scrub jays. Biol Lett. 2012;8(4):582–5. doi: 10.1098/rsbl.2011.1244 . - DOI - PMC - PubMed
-
- Coulon A, Fitzpatrick JW, Bowman R, Lovette IJ. Effects of habitat fragmentation on effective dispersal of Florida scrub-jays. Conserv Biol. 2010;24(4):1080–8. doi: 10.1111/j.1523-1739.2009.01438.x . - DOI - PubMed
-
- Duckworth RA, Badyaev AV. Coupling of dispersal and aggression facilitates the rapid range expansion of a passerine bird. Proc Natl Acad Sci USA. 2007;104(38):15017–22. doi: 10.1073/pnas.0706174104 . - DOI - PMC - PubMed
