

Laminar differences in response to simple and spectro-temporally complex sounds in the primary auditory cortex of ketamine-anesthetized gerbils
- PMID: 28771568
- PMCID: PMC5542772
- DOI: 10.1371/journal.pone.0182514
Laminar differences in response to simple and spectro-temporally complex sounds in the primary auditory cortex of ketamine-anesthetized gerbils
Abstract
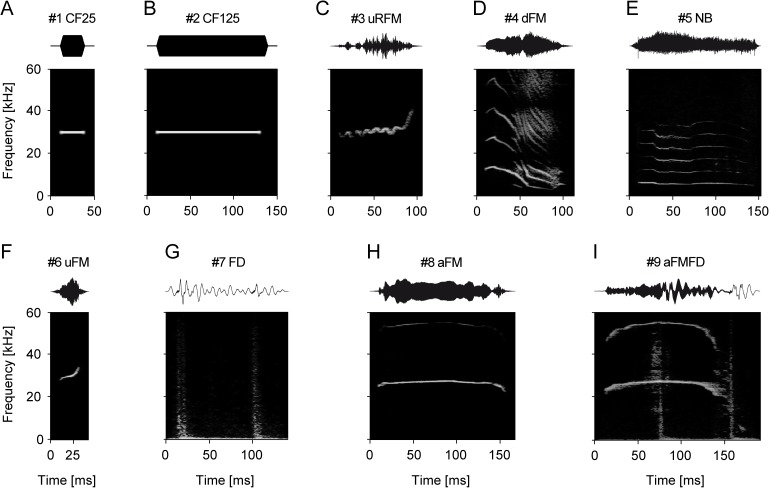
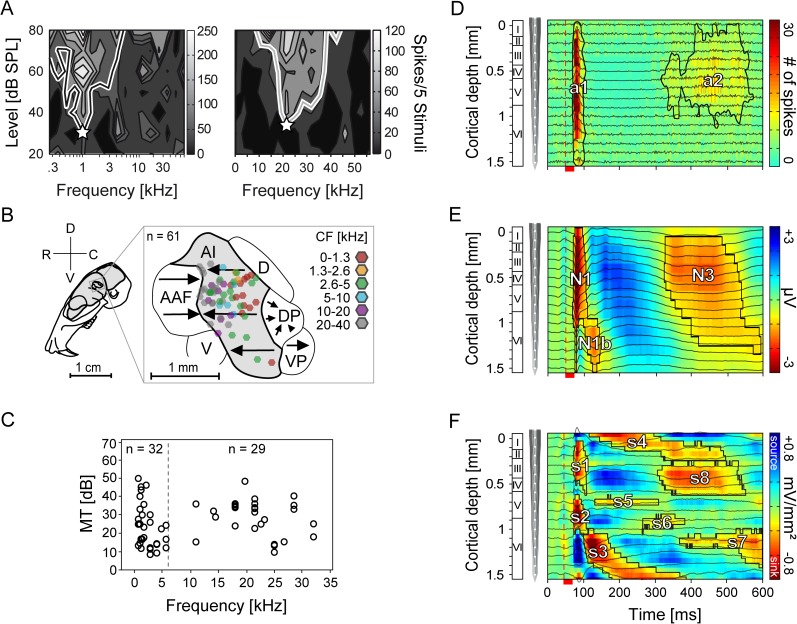
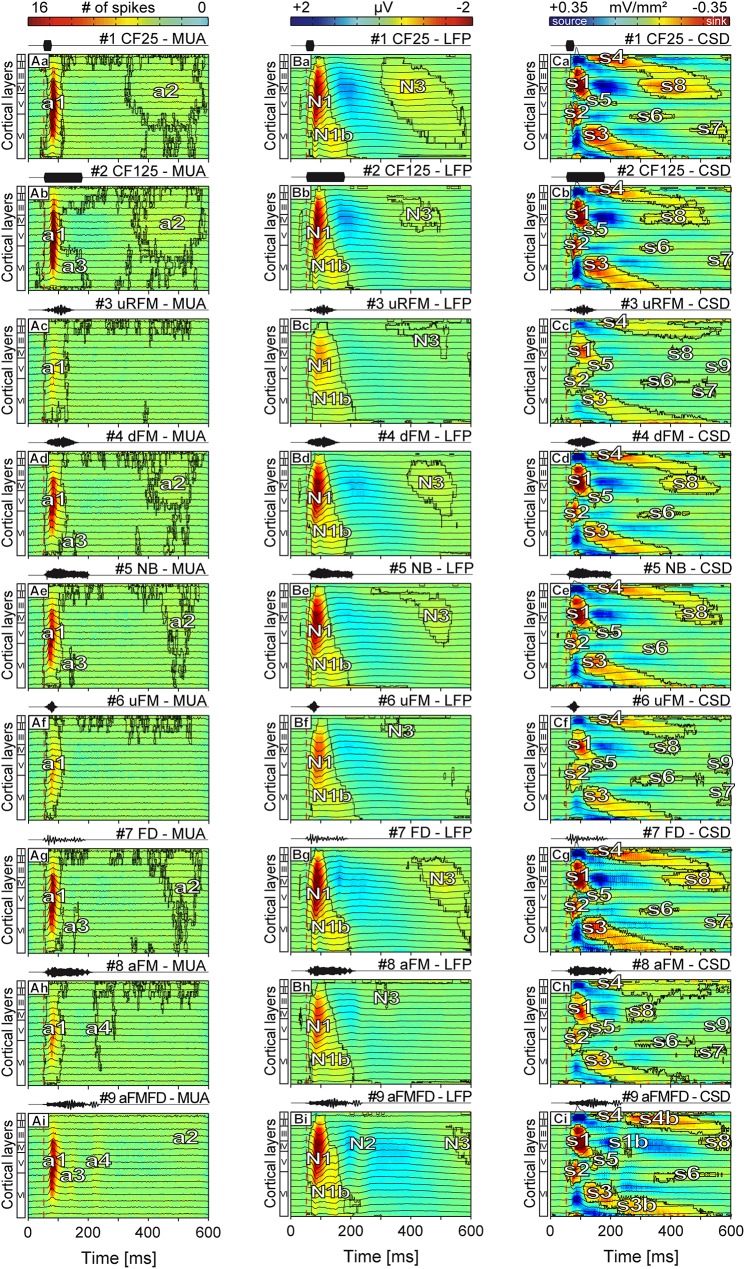
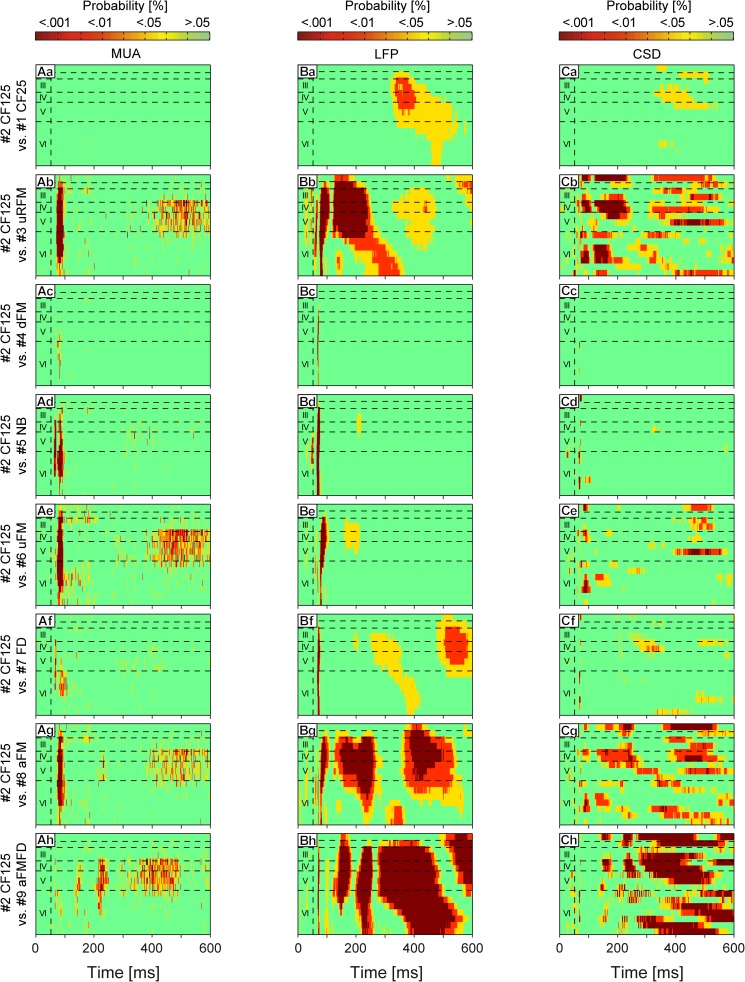
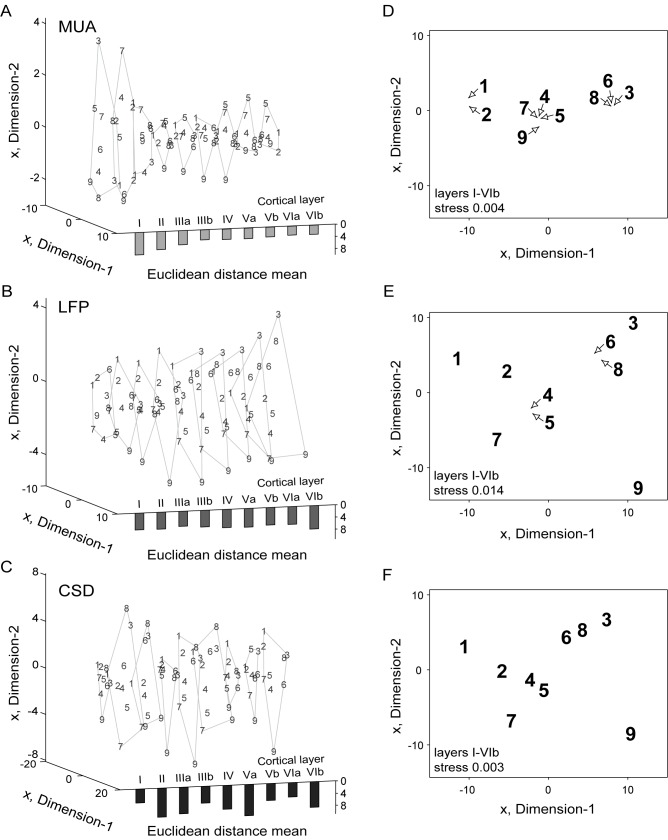
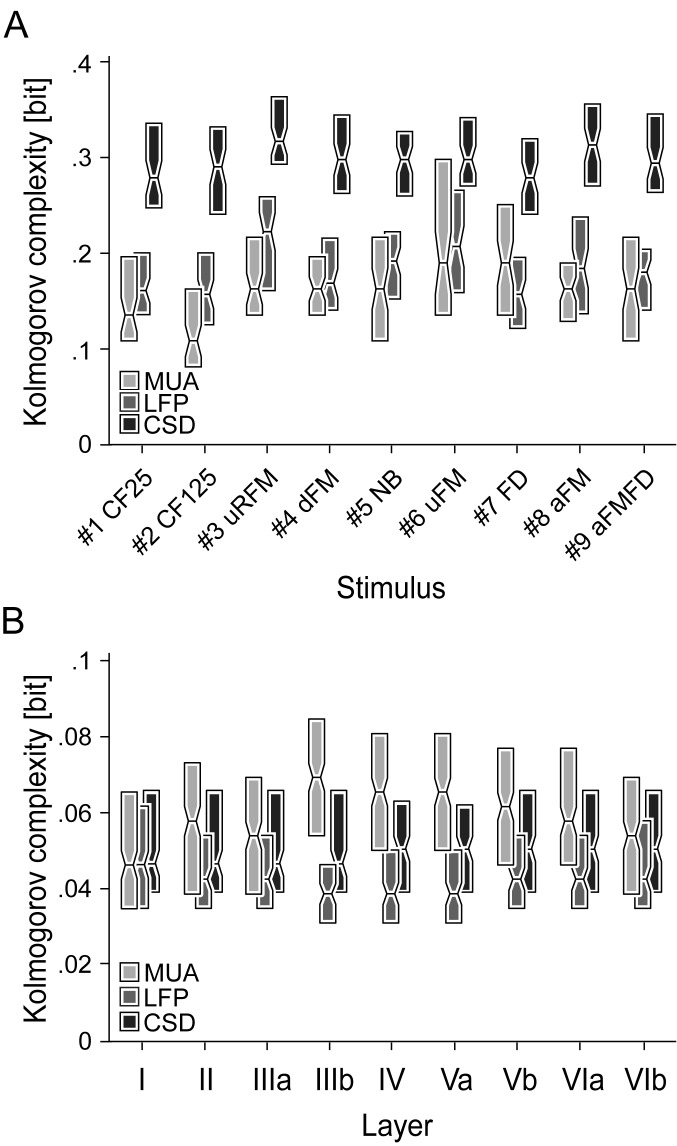
In mammals, acoustic communication plays an important role during social behaviors. Despite their ethological relevance, the mechanisms by which the auditory cortex represents different communication call properties remain elusive. Recent studies have pointed out that communication-sound encoding could be based on discharge patterns of neuronal populations. Following this idea, we investigated whether the activity of local neuronal networks, such as those occurring within individual cortical columns, is sufficient for distinguishing between sounds that differed in their spectro-temporal properties. To accomplish this aim, we analyzed simple pure-tone and complex communication call elicited multi-unit activity (MUA) as well as local field potentials (LFP), and current source density (CSD) waveforms at the single-layer and columnar level from the primary auditory cortex of anesthetized Mongolian gerbils. Multi-dimensional scaling analysis was used to evaluate the degree of "call-specificity" in the evoked activity. The results showed that whole laminar profiles segregated 1.8-2.6 times better across calls than single-layer activity. Also, laminar LFP and CSD profiles segregated better than MUA profiles. Significant differences between CSD profiles evoked by different sounds were more pronounced at mid and late latencies in the granular and infragranular layers and these differences were based on the absence and/or presence of current sinks and on sink timing. The stimulus-specific activity patterns observed within cortical columns suggests that the joint activity of local cortical populations (as local as single columns) could indeed be important for encoding sounds that differ in their acoustic attributes.
Conflict of interest statement
Figures
Similar articles
-
Quantification of mid and late evoked sinks in laminar current source density profiles of columns in the primary auditory cortex.Front Neural Circuits. 2015 Oct 2;9:52. doi: 10.3389/fncir.2015.00052. eCollection 2015. Front Neural Circuits. 2015. PMID: 26557058 Free PMC article.
-
Differential effects of iontophoretic application of the GABAA-antagonists bicuculline and gabazine on tone-evoked local field potentials in primary auditory cortex: interaction with ketamine anesthesia.Brain Res. 2008 Jul 18;1220:58-69. doi: 10.1016/j.brainres.2007.10.023. Epub 2007 Oct 22. Brain Res. 2008. PMID: 18022607
-
The laminar and temporal structure of stimulus information in the phase of field potentials of auditory cortex.J Neurosci. 2011 Nov 2;31(44):15787-801. doi: 10.1523/JNEUROSCI.1416-11.2011. J Neurosci. 2011. PMID: 22049422 Free PMC article.
-
How do auditory cortex neurons represent communication sounds?Hear Res. 2013 Nov;305:102-12. doi: 10.1016/j.heares.2013.03.011. Epub 2013 Apr 17. Hear Res. 2013. PMID: 23603138 Review.
-
Anisotropy of ongoing neural activity in the primate visual cortex.Eye Brain. 2014 Sep 23;6(Suppl 1):113-120. doi: 10.2147/EB.S51822. eCollection 2014. Eye Brain. 2014. PMID: 28539791 Free PMC article. Review.
Cited by
-
Recent advances in understanding the auditory cortex.F1000Res. 2018 Sep 26;7:F1000 Faculty Rev-1555. doi: 10.12688/f1000research.15580.1. eCollection 2018. F1000Res. 2018. PMID: 30345008 Free PMC article. Review.
-
Receptive-field nonlinearities in primary auditory cortex: a comparative perspective.Cereb Cortex. 2024 Sep 3;34(9):bhae364. doi: 10.1093/cercor/bhae364. Cereb Cortex. 2024. PMID: 39270676 Free PMC article.
References
-
- Winter P, Funkenstein HH. The effect of species-specific vocalization on the discharge of auditory cortical cells in the awake squirrel monkey. (Saimiri sciureus). Exp Brain Res. 1973;18(5):489–504. Epub 1973/12/20. . - PubMed
-
- Rauschecker JP. Cortical processing of complex sounds. Curr Opin Neurobiol. 1998;8(4):516–21. Epub 1998/09/30. . - PubMed
-
- Rauschecker JP. Processing of complex sounds in the auditory cortex of cat, monkey, and man. Acta Otolaryngol Suppl. 1997;532:34–8. Epub 1997/01/01. . - PubMed
-
- Kanwal JS, Rauschecker JP. Auditory cortex of bats and primates: managing species-specific calls for social communication. Frontiers in Bioscience. 2007;12:4621–40. doi: 10.2741/2413 - DOI - PMC - PubMed
-
- Steinschneider M, Arezzo J, Vaughan HG Jr. Speech evoked activity in the auditory radiations and cortex of the awake monkey. Brain Res. 1982;252(2):353–65. Epub 1982/12/09. . - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
