

Cellular Timekeeping: It's Redox o'Clock
- PMID: 28778867
- PMCID: PMC5932581
- DOI: 10.1101/cshperspect.a027698
Cellular Timekeeping: It's Redox o'Clock
Abstract
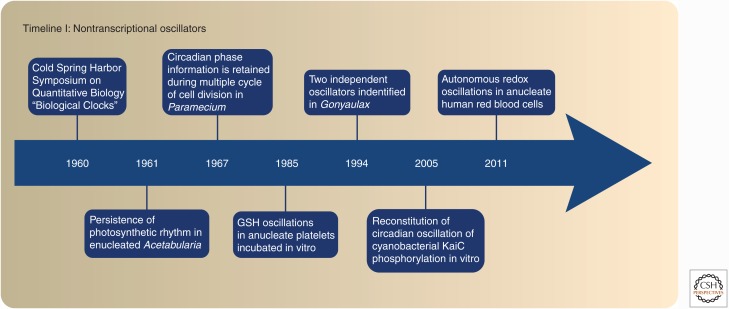
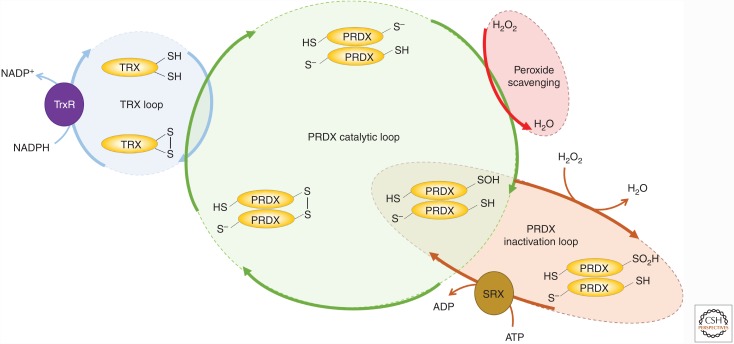
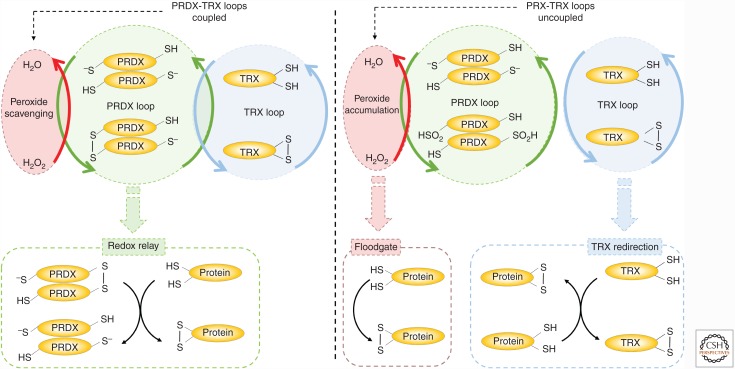
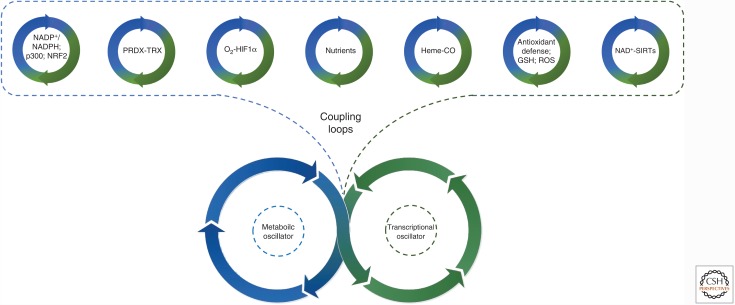
Mounting evidence in recent years supports the extensive interaction between the circadian and redox systems. The existence of such a relationship is not surprising because most organisms, be they diurnal or nocturnal, display daily oscillations in energy intake, locomotor activity, and exposure to exogenous and internally generated oxidants. The transcriptional clock controls the levels of many antioxidant proteins and redox-active cofactors, and, conversely, the cellular redox poise has been shown to feed back to the transcriptional oscillator via redox-sensitive transcription factors and enzymes. However, the circadian cycles in the S-sulfinylation of the peroxiredoxin (PRDX) proteins constituted the first example of an autonomous circadian redox oscillation, which occurred independently of the transcriptional clock. Importantly, the high phylogenetic conservation of these rhythms suggests that they might predate the evolution of the transcriptional oscillator, and therefore could be a part of a primordial circadian redox/metabolic oscillator. This discovery forced the reappraisal of the dogmatic transcription-centered view of the clockwork and opened a new avenue of research. Indeed, the investigation into the links between the circadian and redox systems is still in its infancy, and many important questions remain to be addressed.
Copyright © 2018 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
Similar articles
-
Interplay between cellular redox oscillations and circadian clocks.Diabetes Obes Metab. 2015 Sep;17 Suppl 1:55-64. doi: 10.1111/dom.12519. Diabetes Obes Metab. 2015. PMID: 26332969 Review.
-
Cross-talk between circadian clocks, sleep-wake cycles, and metabolic networks: Dispelling the darkness.Bioessays. 2016 Apr;38(4):394-405. doi: 10.1002/bies.201500056. Epub 2016 Feb 11. Bioessays. 2016. PMID: 26866932 Free PMC article. Review.
-
Circadian redox oscillations and metabolism.Trends Endocrinol Metab. 2015 Aug;26(8):430-7. doi: 10.1016/j.tem.2015.05.012. Epub 2015 Jun 22. Trends Endocrinol Metab. 2015. PMID: 26113283 Free PMC article. Review.
-
Metabolic and nontranscriptional circadian clocks: eukaryotes.Annu Rev Biochem. 2014;83:165-89. doi: 10.1146/annurev-biochem-060713-035623. Epub 2014 Mar 3. Annu Rev Biochem. 2014. PMID: 24606143 Free PMC article. Review.
-
Rethinking the clockwork: redox cycles and non-transcriptional control of circadian rhythms.Biochem Soc Trans. 2014 Feb;42(1):1-10. doi: 10.1042/BST20130169. Biochem Soc Trans. 2014. PMID: 24450621
Cited by
-
Quantitative Studies for Cell-Division Cycle Control.Front Physiol. 2019 Aug 19;10:1022. doi: 10.3389/fphys.2019.01022. eCollection 2019. Front Physiol. 2019. PMID: 31496950 Free PMC article. Review.
-
Keeping development on time: Insights into post-transcriptional mechanisms driving oscillatory gene expression during vertebrate segmentation.Wiley Interdiscip Rev RNA. 2023 Jan;14(1):e1751. doi: 10.1002/wrna.1751. Epub 2022 Jul 19. Wiley Interdiscip Rev RNA. 2023. PMID: 35851751 Free PMC article. Review.
-
Uncoupled redox stress: how a temporal misalignment of redox-regulated processes and circadian rhythmicity exacerbates the stressed state.Open Biol. 2023 Sep;13(9):230151. doi: 10.1098/rsob.230151. Epub 2023 Sep 6. Open Biol. 2023. PMID: 37669692 Free PMC article. Review.
-
Longevity strategies in response to light in the reef coral Stylophora pistillata.Sci Rep. 2020 Nov 17;10(1):19937. doi: 10.1038/s41598-020-76925-2. Sci Rep. 2020. PMID: 33203910 Free PMC article.
-
Role of the Circadian Gas-Responsive Hemeprotein NPAS2 in Physiology and Pathology.Biology (Basel). 2023 Oct 22;12(10):1354. doi: 10.3390/biology12101354. Biology (Basel). 2023. PMID: 37887064 Free PMC article. Review.
References
-
- Adamovich Y, Ladeuix B, Golik M, Koeners MP, Asher G. 2016. Rhythmic oxygen levels reset circadian clocks through HIF1α. Cell Metab 25: 93–101. - PubMed
-
- Asher G, Gatfield D, Stratmann M, Reinke H, Dibner C, Kreppel F, Mostoslavsky R, Alt FW, Schibler U. 2008. SIRT1 regulates circadian clock gene expression through PER2 deacetylation. Cell 134: 317–328. - PubMed
-
- Asher G, Reinke H, Altmeyer M, Gutierrez-Arcelus M, Hottiger MO, Schibler U. 2010. Poly(ADP-ribose) polymerase 1 participates in the phase entrainment of circadian clocks to feeding. Cell 142: 943–953. - PubMed
-
- Avitabile D, Nicolussi A, Capriotti AL, Cucina A, Samperi R, Bizzarri M, Torrisi MR. 2014. Peroxiredoxin 2 nuclear levels are regulated by circadian clock synchronization in human keratinocytes. Int J Biochem Cell Biol 53: 24–34. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources