

A Commensal Bifidobacterium longum Strain Prevents Gluten-Related Immunopathology in Mice through Expression of a Serine Protease Inhibitor
- PMID: 28778891
- PMCID: PMC5601352
- DOI: 10.1128/AEM.01323-17
A Commensal Bifidobacterium longum Strain Prevents Gluten-Related Immunopathology in Mice through Expression of a Serine Protease Inhibitor
Abstract
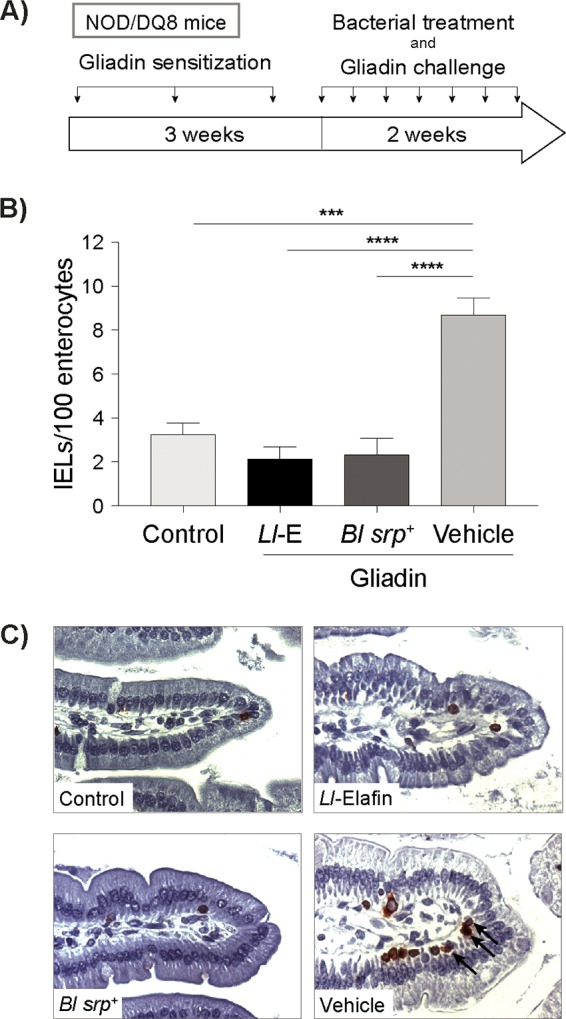
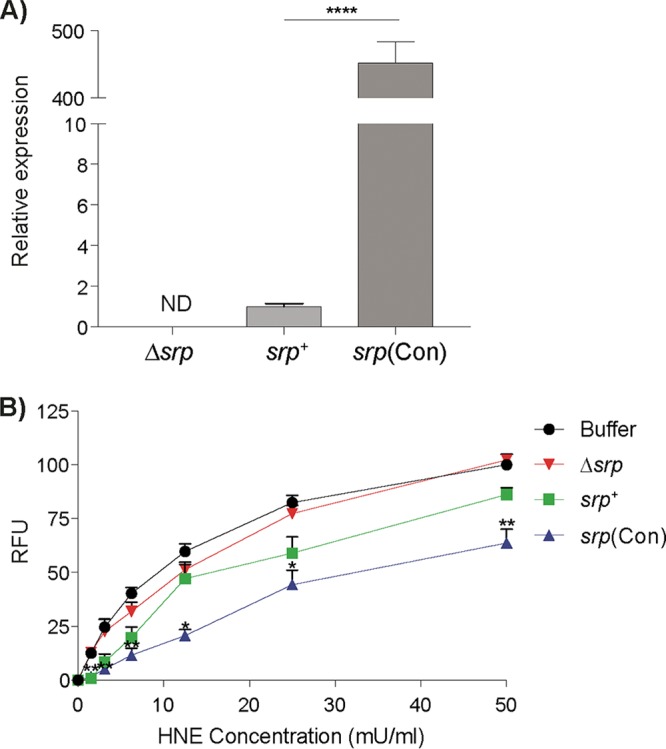
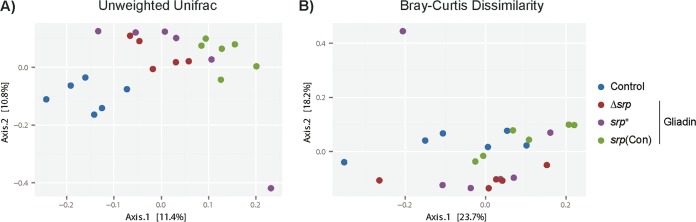
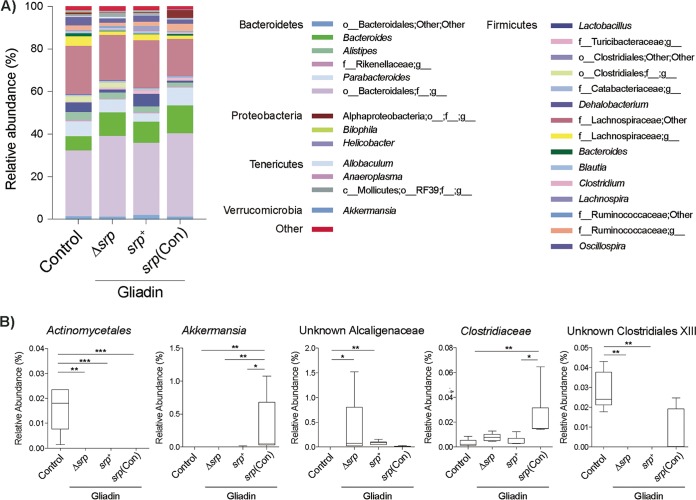
Microbiota-modulating strategies, including probiotic administration, have been tested for the treatment of chronic gastrointestinal diseases despite limited information regarding their mechanisms of action. We previously demonstrated that patients with active celiac disease have decreased duodenal expression of elafin, a human serine protease inhibitor, and supplementation of elafin by a recombinant Lactococcus lactis strain prevents gliadin-induced immunopathology in the NOD/DQ8 mouse model of gluten sensitivity. The commensal probiotic strain Bifidobacterium longum NCC2705 produces a serine protease inhibitor (Srp) that exhibits immune-modulating properties. Here, we demonstrate that B. longum NCC2705, but not a srp knockout mutant, attenuates gliadin-induced immunopathology and impacts intestinal microbial composition in NOD/DQ8 mice. Our results highlight the beneficial effects of a serine protease inhibitor produced by commensal B. longum strains.IMPORTANCE Probiotic therapies have been widely used to treat gastrointestinal disorders with variable success and poor mechanistic insight. Delivery of specific anti-inflammatory molecules has been limited to the use of genetically modified organisms, which has raised some public and regulatory concerns. By examining a specific microbial product naturally expressed by a commensal bacterial strain, we provide insight into a mechanistic basis for the use of B. longum NCC2705 to help treat gluten-related disorders.
Keywords: celiac; commensal; gluten; microbiota; probiotic; serpin.
Copyright © 2017 American Society for Microbiology.
Figures

Similar articles
-
Novel role of the serine protease inhibitor elafin in gluten-related disorders.Am J Gastroenterol. 2014 May;109(5):748-56. doi: 10.1038/ajg.2014.48. Epub 2014 Apr 8. Am J Gastroenterol. 2014. PMID: 24710505 Free PMC article.
-
Intestinal microbiota modulates gluten-induced immunopathology in humanized mice.Am J Pathol. 2015 Nov;185(11):2969-82. doi: 10.1016/j.ajpath.2015.07.018. Epub 2015 Oct 9. Am J Pathol. 2015. PMID: 26456581 Free PMC article.
-
Carbohydrate-controlled serine protease inhibitor (serpin) production in Bifidobacterium longum subsp. longum.Sci Rep. 2021 Mar 31;11(1):7236. doi: 10.1038/s41598-021-86740-y. Sci Rep. 2021. PMID: 33790385 Free PMC article.
-
Bosom Buddies: The Symbiotic Relationship Between Infants and Bifidobacterium longum ssp. longum and ssp. infantis. Genetic and Probiotic Features.Annu Rev Food Sci Technol. 2016;7:1-21. doi: 10.1146/annurev-food-041715-033151. Annu Rev Food Sci Technol. 2016. PMID: 26934170 Review.
-
Celiac disease: risk assessment, diagnosis, and monitoring.Mol Diagn Ther. 2008;12(5):289-98. doi: 10.1007/BF03256294. Mol Diagn Ther. 2008. PMID: 18803427 Review.
Cited by
-
Identification of a prototype human gut Bifidobacterium longum subsp. longum strain based on comparative and functional genomic approaches.Front Microbiol. 2023 Feb 8;14:1130592. doi: 10.3389/fmicb.2023.1130592. eCollection 2023. Front Microbiol. 2023. PMID: 36846784 Free PMC article.
-
Adherence to Gluten-Free Diet Restores Alpha Diversity in Celiac People but the Microbiome Composition Is Different to Healthy People.Nutrients. 2022 Jun 14;14(12):2452. doi: 10.3390/nu14122452. Nutrients. 2022. PMID: 35745182 Free PMC article.
-
Gut Feelings: How Microbes, Diet, and Host Immunity Shape Disease.Biomedicines. 2025 May 31;13(6):1357. doi: 10.3390/biomedicines13061357. Biomedicines. 2025. PMID: 40564077 Free PMC article. Review.
-
Bacteria: Potential Make-or-Break Determinants of Celiac Disease.Int J Mol Sci. 2024 Feb 8;25(4):2090. doi: 10.3390/ijms25042090. Int J Mol Sci. 2024. PMID: 38396767 Free PMC article. Review.
-
The Gut Microbiota in Celiac Disease and probiotics.Nutrients. 2019 Oct 5;11(10):2375. doi: 10.3390/nu11102375. Nutrients. 2019. PMID: 31590358 Free PMC article. Review.
References
-
- Pinto-Sanchez MI, Smecuol EC, Temprano MP, Sugai E, Gonzalez A, Moreno ML, Huang X, Bercik P, Cabanne A, Vazquez H, Niveloni S, Mazure R, Maurino E, Verdu EF, Bai JC. 15 September 2016. Bifidobacterium infantis NLS super strain reduces the expression of alpha-defensin-5, a marker of innate immunity, in the mucosa of active celiac disease patients. J Clin Gastroenterol doi:10.1097/MCG.0000000000000687. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
