

Defining a critical period for inhibitory circuits within the somatosensory cortex
- PMID: 28779074
- PMCID: PMC5544762
- DOI: 10.1038/s41598-017-07400-8
Defining a critical period for inhibitory circuits within the somatosensory cortex
Abstract
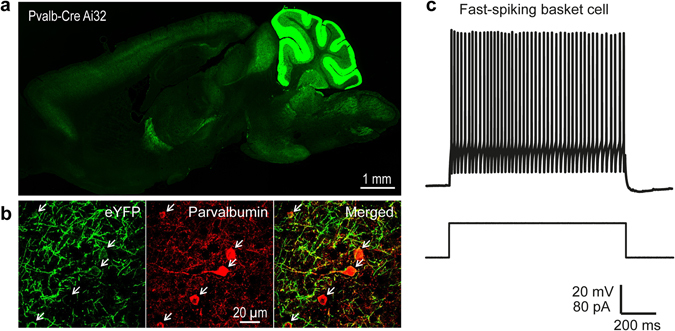
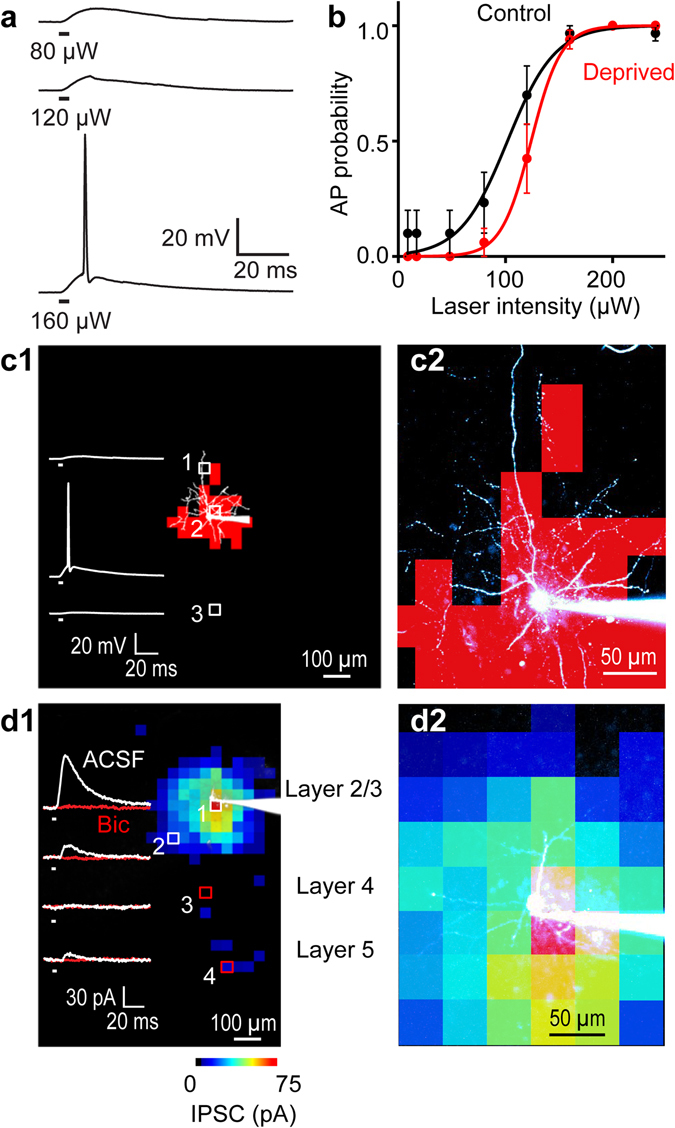
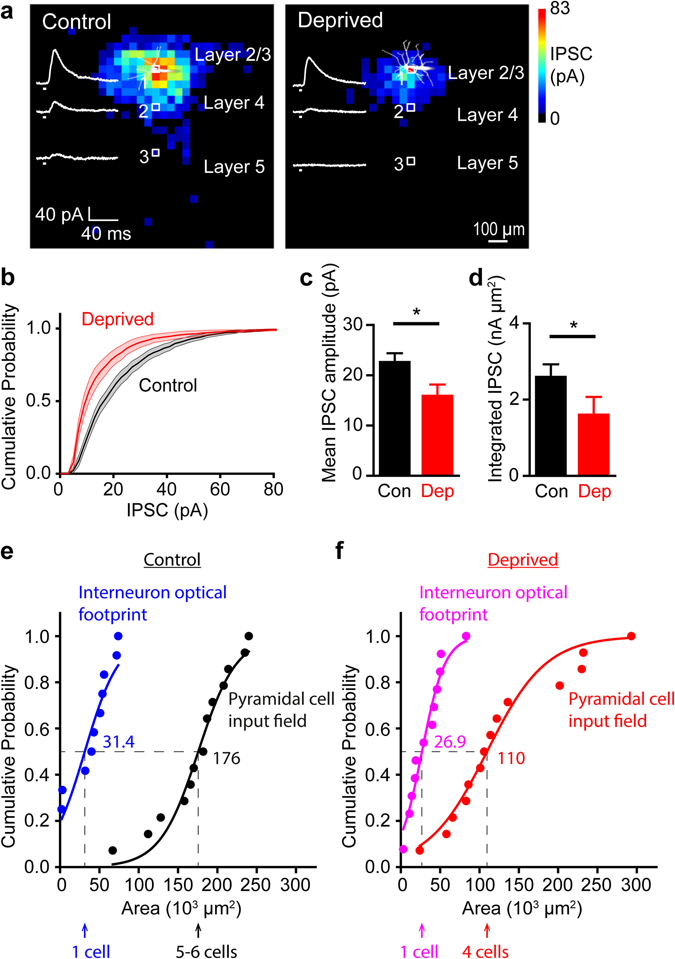
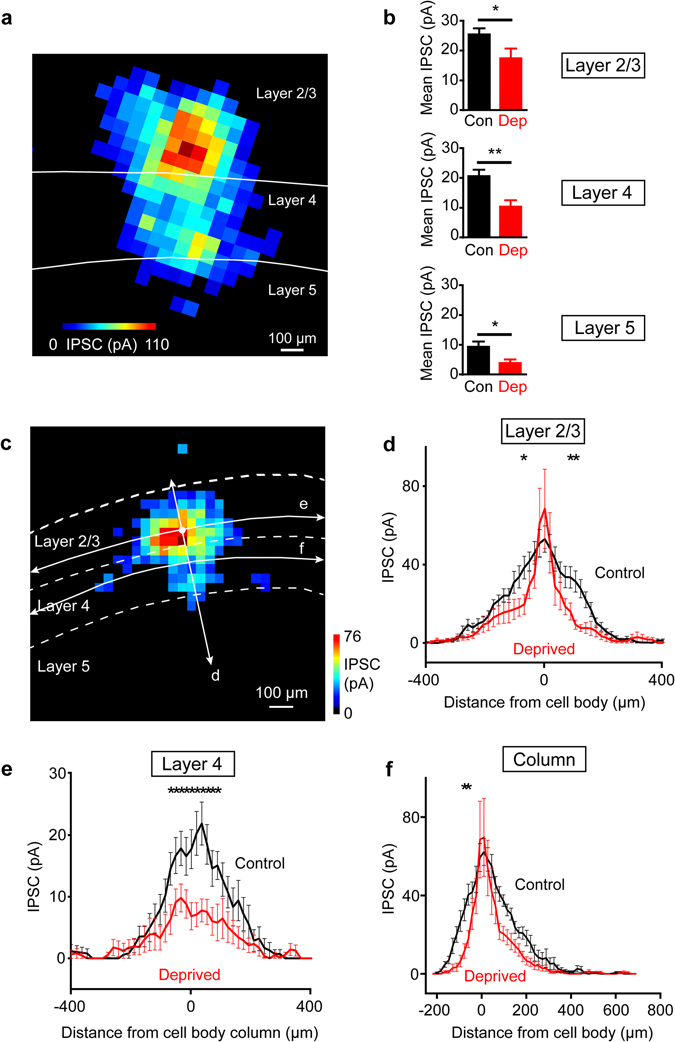
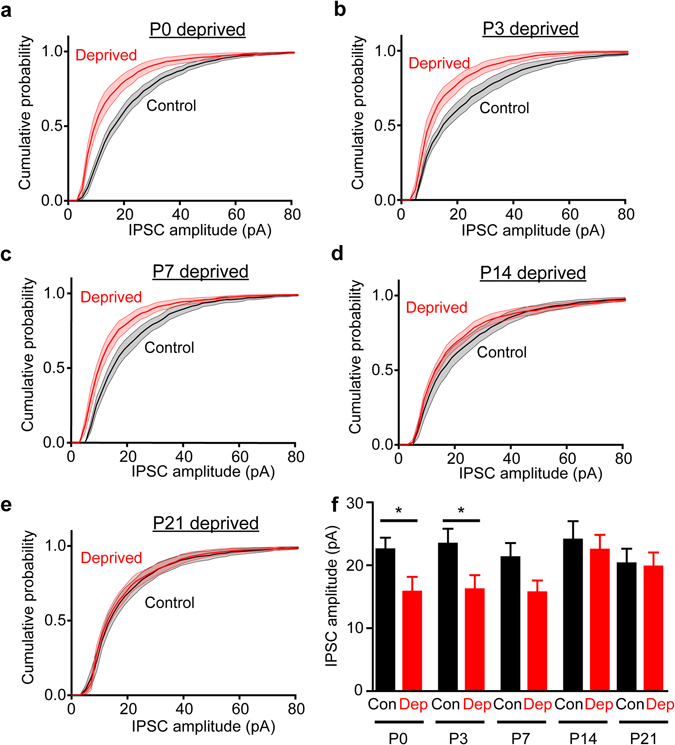
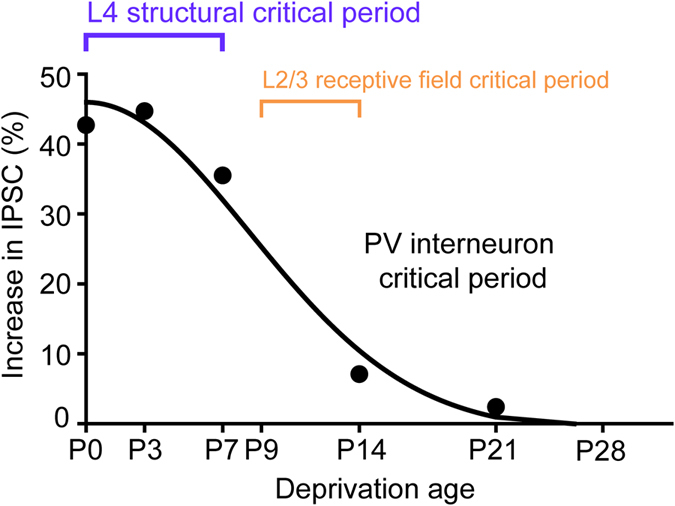
Although experience-dependent changes in brain inhibitory circuits are thought to play a key role during the "critical period" of brain development, the nature and timing of these changes are poorly understood. We examined the role of sensory experience in sculpting an inhibitory circuit in the primary somatosensory cortex (S1) of mice by using optogenetics to map the connections between parvalbumin (PV) expressing interneurons and layer 2/3 pyramidal cells. Unilateral whisker deprivation decreased the strength and spatial range of inhibitory input provided to pyramidal neurons by PV interneurons in layers 2/3, 4 and 5. By varying the time when sensory input was removed, we determined that the critical period closes around postnatal day 14. This yields the first precise time course of critical period plasticity for an inhibitory circuit.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Wiesel TN, Hubel DH. Effects of visual deprivation on morphology and physiology of cells in the cats lateral geniculate body. J Neurophysiol. 1963;26:978–993. - PubMed
-
- Wiesel TN, Hubel DH. Single-cell responses in striate cortex of kittens deprived of vision in one eye. J Neurophysiol. 1963;26:1003–1017. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
