

Robust circadian clock oscillation and osmotic rhythms in inner medulla reflecting cortico-medullary osmotic gradient rhythm in rodent kidney
- PMID: 28779094
- PMCID: PMC5544761
- DOI: 10.1038/s41598-017-07767-8
Robust circadian clock oscillation and osmotic rhythms in inner medulla reflecting cortico-medullary osmotic gradient rhythm in rodent kidney
Abstract
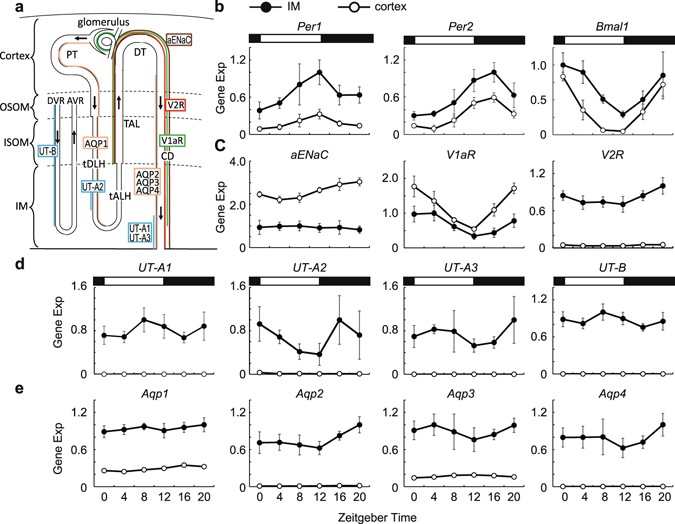
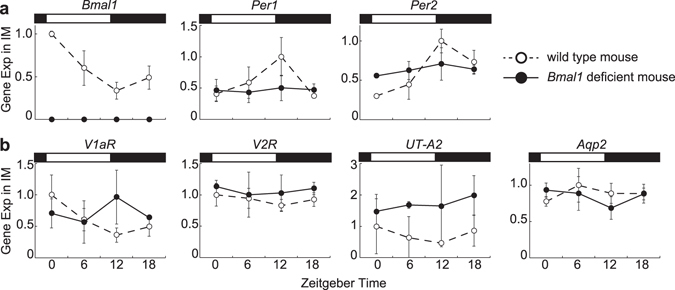
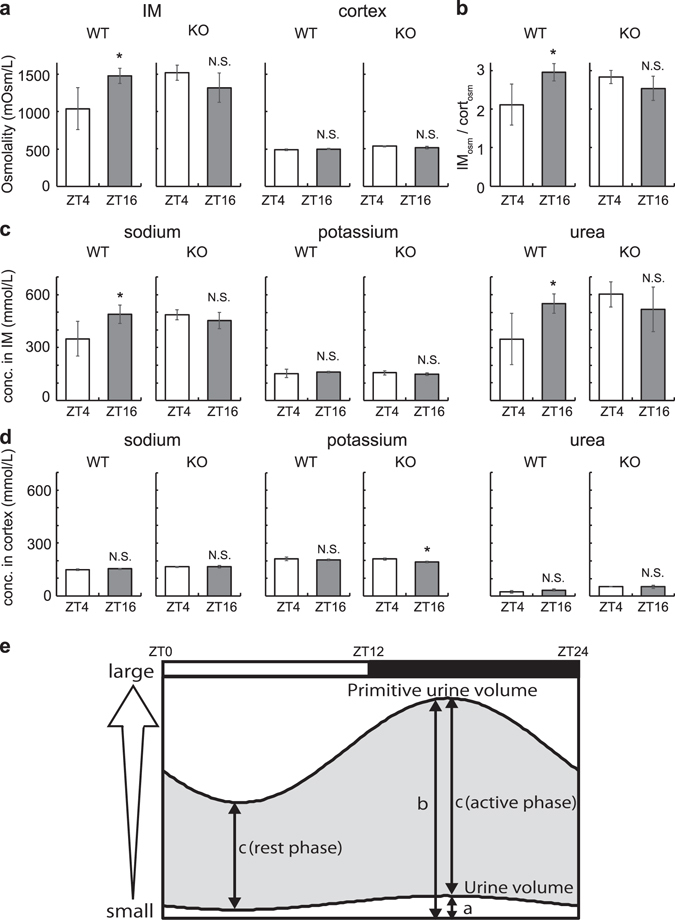
Circadian clocks in mammals function in most organs and tissues throughout the body. Various renal functions such as the glomerular filtration and excretion of electrolytes exhibit circadian rhythms. Although it has been reported that the expression of the clock genes composing molecular oscillators show apparent daily rhythms in rodent kidneys, functional variations of regional clocks are not yet fully understood. In this study, using macroscopic bioluminescence imaging method of the PER2::Luciferase knock-in mouse kidney, we reveal that strong and robust circadian clock oscillation is observed in the medulla. In addition, the osmotic pressure in the inner medulla shows apparent daily fluctuation, but not in the cortex. Quantitative-PCR analysis of the genes contributing to the generation of high osmotic pressure or the water re-absorption in the inner medulla, such as vasopressin receptors (V1aR, V2R), urea transporter (UT-A2) and water channel (Aqp2) show diurnal variations as well as clock genes. Deficiency of an essential clock gene Bmal1 impairs day-night variations of osmotic pressure gradient in the inner medulla, suggesting that circadian clocks in the medulla part of the kidney may regulate the circadian rhythm of cortico-medullary osmotic pressure gradient, and may contribute physiological day-night rhythm of urination.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
