

cAMP receptor protein regulates mouse colonization, motility, fimbria-mediated adhesion, and stress tolerance in uropathogenic Proteus mirabilis
- PMID: 28779108
- PMCID: PMC5544767
- DOI: 10.1038/s41598-017-07304-7
cAMP receptor protein regulates mouse colonization, motility, fimbria-mediated adhesion, and stress tolerance in uropathogenic Proteus mirabilis
Abstract
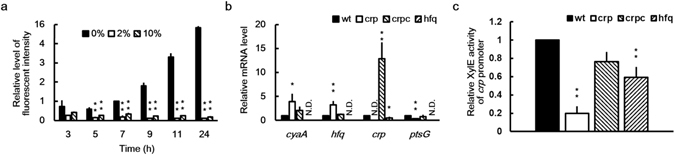
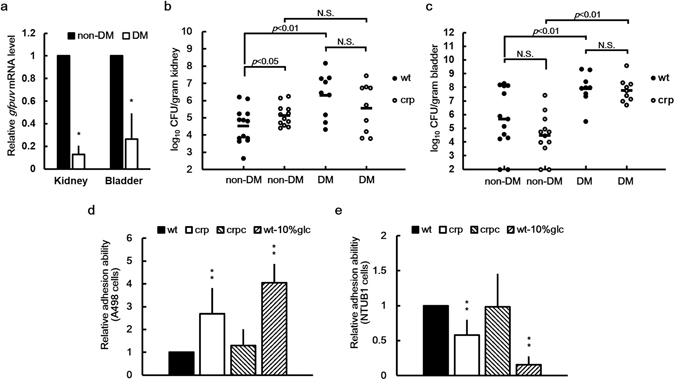
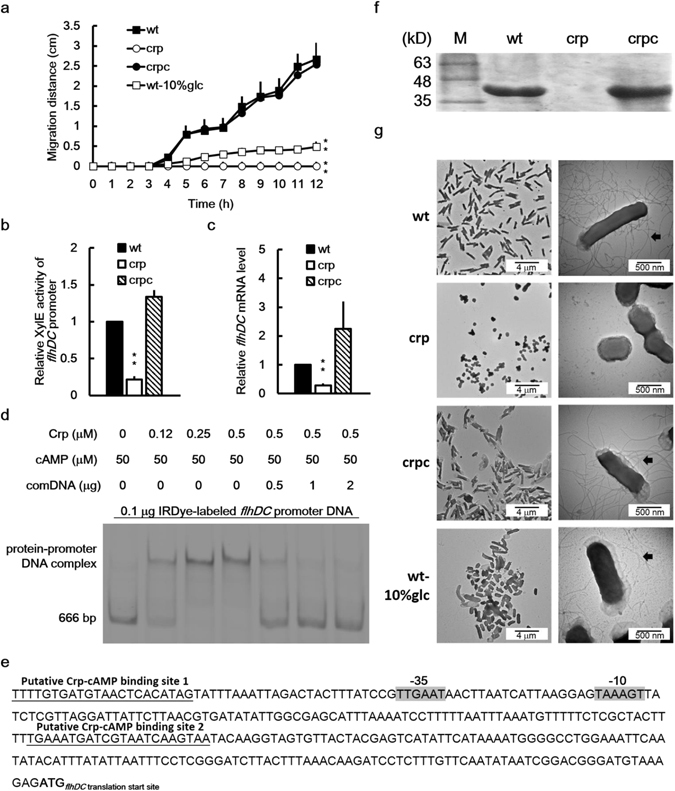
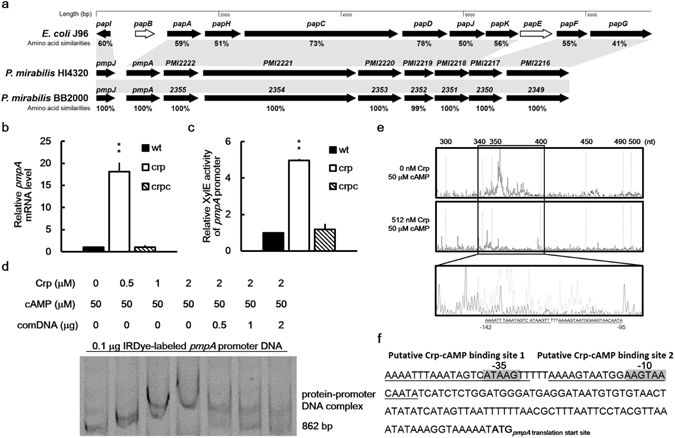
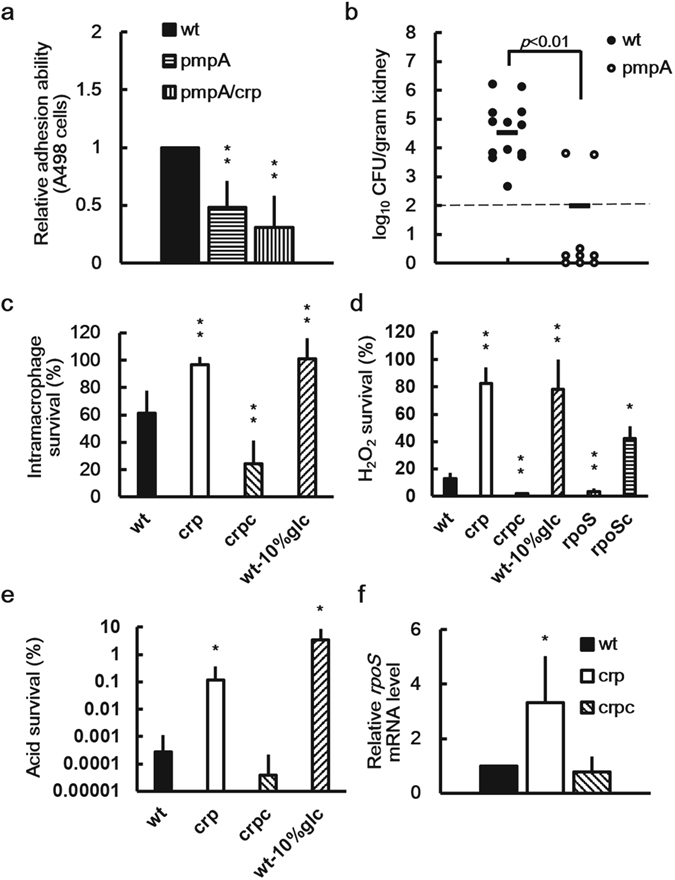
Cyclic AMP receptor protein (Crp) is a major transcriptional regulator in bacteria. This study demonstrated that Crp affects numerous virulence-related phenotypes, including colonization of mice, motility, fimbria-mediated adhesion, and glucose stress tolerance in uropathogenic Proteus mirabilis. Diabetic mice were more susceptible to kidney colonization by wild-type strain than nondiabetic mice, in which the crp mutant exhibited increased kidney colonization. Loss of crp or addition of 10% glucose increased the P. mirabilis adhesion to kidney cells. Direct negative regulation of pmpA (which encodes the major subunit of P-like fimbriae) expression by Crp was demonstrated using a reporter assay and DNase I footprinting. Moreover, the pmpA/crp double mutant exhibited reduced kidney adhesion comparable to that of the pmpA mutant, and mouse kidney colonization by the pmpA mutant was significantly attenuated. Hence, the upregulation of P-like fimbriae in the crp mutant substantially enhanced kidney colonization. Moreover, increased survival in macrophages, increased stress tolerance, RpoS upregulation, and flagellum deficiency leading to immune evasion may promote kidney colonization by the crp mutant. This is the first study to elucidate the role of Crp in the virulence of uropathogenic P. mirabilis, underlying mechanisms, and related therapeutic potential.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
