

Excessive alcohol consumption induces methane production in humans and rats
- PMID: 28779149
- PMCID: PMC5544731
- DOI: 10.1038/s41598-017-07637-3
Excessive alcohol consumption induces methane production in humans and rats
Abstract
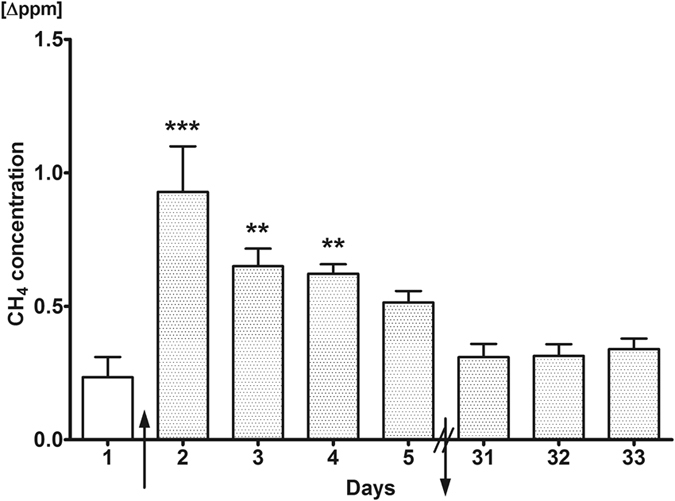
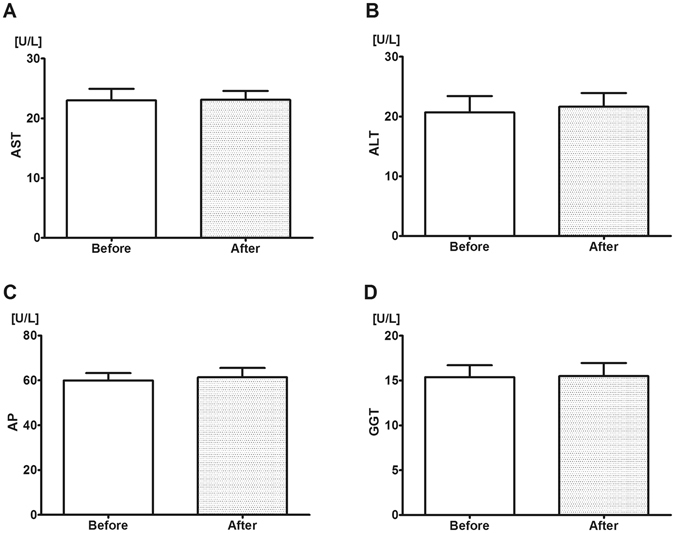
Various studies have established the possibility of non-bacterial methane (CH4) generation in oxido-reductive stress conditions in plants and animals. Increased ethanol input is leading to oxido-reductive imbalance in eukaryotes, thus our aim was to provide evidence for the possibility of ethanol-induced methanogenesis in non-CH4 producer humans, and to corroborate the in vivo relevance of this pathway in rodents. Healthy volunteers consumed 1.15 g/kg/day alcohol for 4 days and the amount of exhaled CH4 was recorded by high sensitivity photoacoustic spectroscopy. Additionally, Sprague-Dawley rats were allocated into control, 1.15 g/kg/day and 2.7 g/kg/day ethanol-consuming groups to detect the whole-body CH4 emissions and mitochondrial functions in liver and hippocampus samples with high-resolution respirometry. Mitochondria-targeted L-alpha-glycerylphosphorylcholine (GPC) can increase tolerance to liver injury, thus the effects of GPC supplementations were tested in further ethanol-fed groups. Alcohol consumption was accompanied by significant CH4 emissions in both human and rat series of experiments. 2.7 g/kg/day ethanol feeding reduced the oxidative phosphorylation capacity of rat liver mitochondria, while GPC significantly decreased the alcohol-induced CH4 formation and hepatic mitochondrial dysfunction as well. These data demonstrate a potential for ethanol to influence human methanogenesis, and suggest a biomarker role for exhaled CH4 in association with mitochondrial dysfunction.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
