

LIN-32/Atonal Controls Oxygen Sensing Neuron Development in Caenorhabditis elegans
- PMID: 28779171
- PMCID: PMC5544745
- DOI: 10.1038/s41598-017-07876-4
LIN-32/Atonal Controls Oxygen Sensing Neuron Development in Caenorhabditis elegans
Abstract
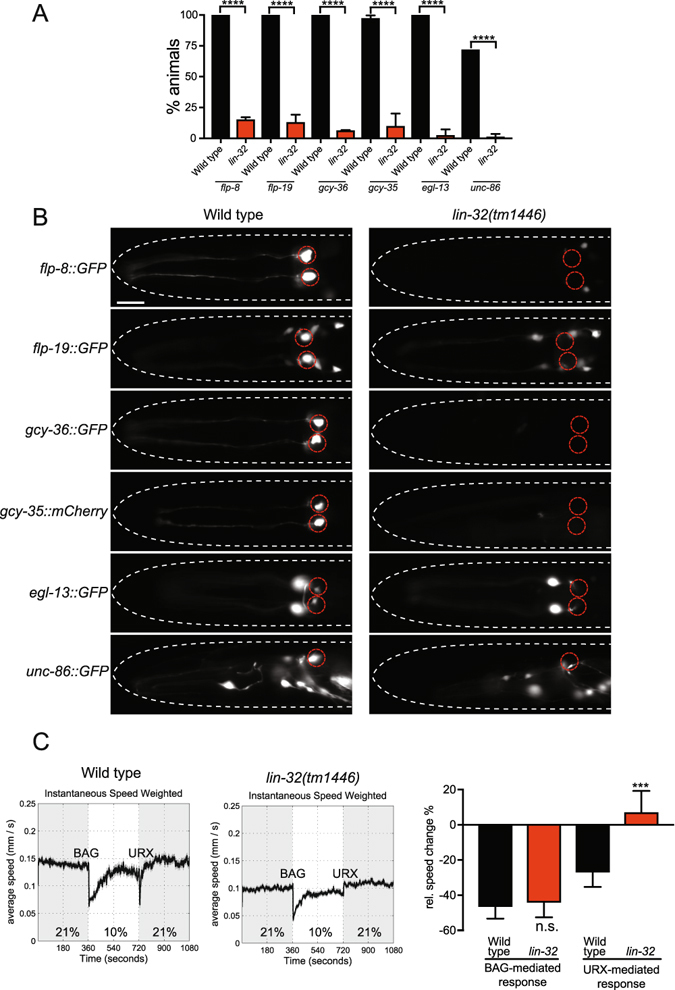
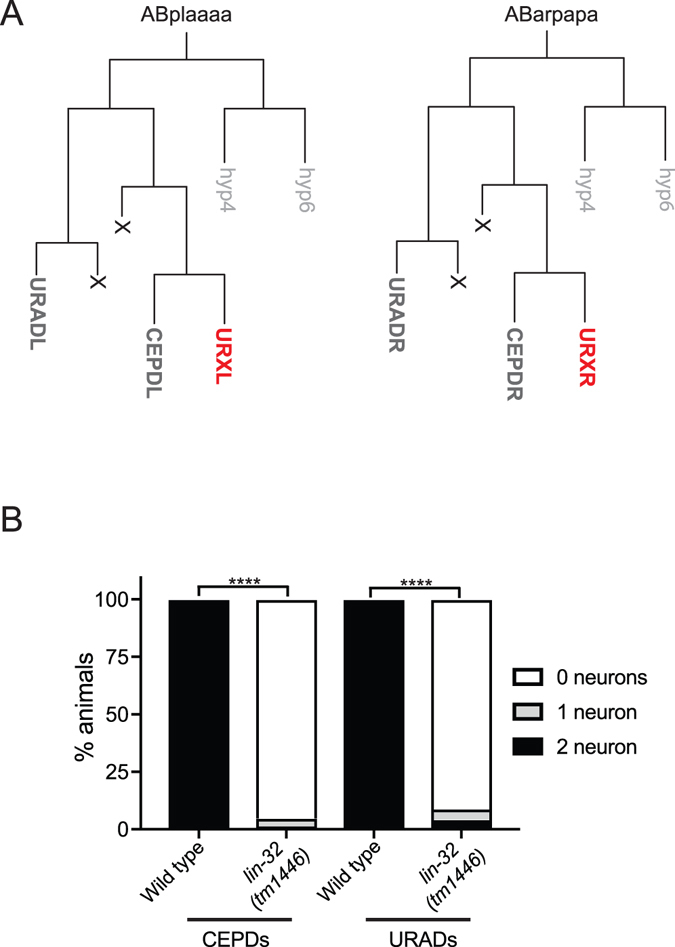
Development of complex nervous systems requires precisely controlled neurogenesis. The generation and specification of neurons occur through the transcriptional and post-transcriptional control of complex regulatory networks. In vertebrates and invertebrates, the proneural basic-helix-loop-helix (bHLH) family of transcription factors has multiple functions in neurogenesis. Here, we identified the LIN-32/Atonal bHLH transcription factor as a key regulator of URXL/R oxygen-sensing neuron development in Caenorhabditis elegans. When LIN-32/Atonal expression is lost, the expression of URX specification and terminal differentiation genes is abrogated. As such, lin-32 mutant animals are unable to respond to increases in environmental oxygen. The URX neurons are generated from a branch of the cell lineage that also produces the CEPDL/R and URADL/R neurons. We found development of these neurons is also defective, suggesting that LIN-32/Atonal regulates neuronal development of the entire lineage. Finally, our results show that aspects of URX neuronal fate are partially restored in lin-32 mutant animals when the apoptosis pathway is inhibited. This suggests that, as in other organisms, LIN-32/Atonal regulates neuronal apoptosis.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures



Similar articles
-
Neurogenesis in Caenorhabditis elegans.Genetics. 2024 Oct 7;228(2):iyae116. doi: 10.1093/genetics/iyae116. Genetics. 2024. PMID: 39167071 Free PMC article. Review.
-
Piecemeal regulation of convergent neuronal lineages by bHLH transcription factors in Caenorhabditis elegans.Development. 2021 Jun 1;148(11):dev199224. doi: 10.1242/dev.199224. Epub 2021 Jun 8. Development. 2021. PMID: 34100067 Free PMC article.
-
The basic helix-loop-helix transcription factors LIN-32 and HLH-2 function together in multiple steps of a C. elegans neuronal sublineage.Development. 2000 Dec;127(24):5415-26. doi: 10.1242/dev.127.24.5415. Development. 2000. PMID: 11076762
-
EGL-13/SoxD specifies distinct O2 and CO2 sensory neuron fates in Caenorhabditis elegans.PLoS Genet. 2013 May;9(5):e1003511. doi: 10.1371/journal.pgen.1003511. Epub 2013 May 9. PLoS Genet. 2013. PMID: 23671427 Free PMC article.
-
A map of terminal regulators of neuronal identity in Caenorhabditis elegans.Wiley Interdiscip Rev Dev Biol. 2016 Jul;5(4):474-98. doi: 10.1002/wdev.233. Epub 2016 May 2. Wiley Interdiscip Rev Dev Biol. 2016. PMID: 27136279 Free PMC article. Review.
Cited by
-
Regulation of Gliogenesis by lin-32/Atoh1 in Caenorhabditis elegans.G3 (Bethesda). 2020 Sep 2;10(9):3271-3278. doi: 10.1534/g3.120.401547. G3 (Bethesda). 2020. PMID: 32665354 Free PMC article.
-
Mechanisms of lineage specification in Caenorhabditis elegans.Genetics. 2023 Dec 6;225(4):iyad174. doi: 10.1093/genetics/iyad174. Genetics. 2023. PMID: 37847877 Free PMC article.
-
Neurogenesis in Caenorhabditis elegans.Genetics. 2024 Oct 7;228(2):iyae116. doi: 10.1093/genetics/iyae116. Genetics. 2024. PMID: 39167071 Free PMC article. Review.
-
Lineage-specific control of convergent differentiation by a Forkhead repressor.Development. 2021 Oct 1;148(19):dev199493. doi: 10.1242/dev.199493. Epub 2021 Sep 28. Development. 2021. PMID: 34423346 Free PMC article.
-
Piecemeal regulation of convergent neuronal lineages by bHLH transcription factors in Caenorhabditis elegans.Development. 2021 Jun 1;148(11):dev199224. doi: 10.1242/dev.199224. Epub 2021 Jun 8. Development. 2021. PMID: 34100067 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
